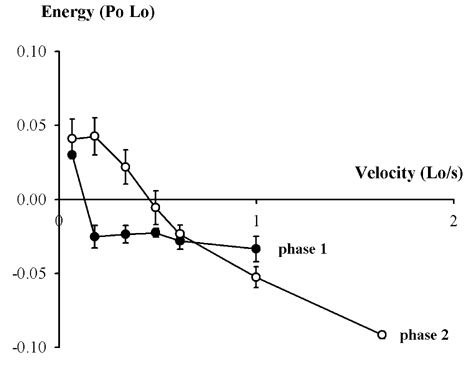
Experiments on whole muscles from amphibians (Hill & Howarth, 1959) and fibre bundles from mouse (Constable et al. 1997) show that muscle can absorb more energy than it releases during stretch. We have reinvestigated this phenomenon with a view to discovering the fate of this energy. Experiments were done on single fibres from frog. Donor animals were humanely killed. Heat production was measured with a thermopile designed for improved spatial resolution. The fibres were mounted between a motor and a force transducer. Constant velocity stretches were applied at the plateau of the isometric tetanus. Work was calculated from force and length change. Heat was evaluated from temperature change recorded by the thermopile. As we have previously reported (Woledge et al. 2000) there is net energy (work + heat) absorption during the stretch when the velocity of stretch is moderately fast, greater than 0.5 Lo s-1. (Lo is the fibre length at which isometric force is maximum.) We report here that energy absorption occurred in two phases. In the initial part of the stretch the rate of energy absorption is greater than in the later part. Records from stretches at different velocities showed that only a limited amount of energy is absorbed in the initial phase (see Fig. 1). In the later phase energy absorption continues for the entire period of stretch without reaching a limit and more energy is absorbed the faster the velocity of stretch. The results suggest that two distinct processes absorb energy. (a) The initial process may be the transition from the ‘isometric distribution’ of cross-bridge states to the ‘stretch distribution’. Once the cross-bridges are in the steady-state ‘stretch distribution’, they cannot store more energy; they behave like brakes converting all of the work done on them into heat. (b) The other energy absorbing process may be the stretching of elastic structures in parallel with cross-bridges (Morgan, 1994; Edman & Tsuchiya, 1996).
This work was funded by The Wellcome Trust.