If breathing is to be established at birth then the liquid secreted into the lung lumen during fetal life must first be removed from the potential air spaces. This occurs during the final stages of pregnancy and is driven by the β-adrenoceptor-mediated transport of Na+ out of the alveolar space (Olver et al. 1986). It has been suggested that this stimulation of Na+ transport involves the insertion of additional Na+ channels into the apical plasma membrane by an exocytotic mechanism that is sensitive to brefeldin A (Ito et al. 1997). Recent work from this laboratory has shown that β-adrenoceptor agonists do increase GNa in FDLE cells (Collett et al. 2002) and so we now explore the effects of brefeldin-A upon this response.
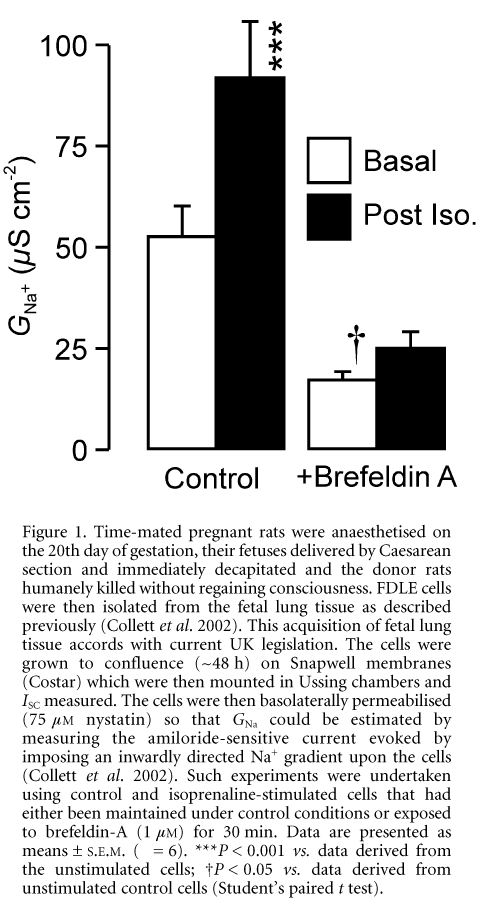
These data confirm (Collett et al. 2002) that isoprenaline-evoked increase in ISC (basal, 7.9 ± 0.7 mA cm-2; post-isoprenaline, 11.2 ± 0.9 mA cm-2, P < 0.02) is normally associated with a rise in GNa (Fig. 1). Exposing unstimulated cells to brefeldin-A (1 mM) caused a barely discernible rise in ISC that developed over a period of ~5 min. The current then declined slowly, reaching a value that was 88.7 ± 3.6 % of its initial level (P < 0.05) after 30 min. Application of isoprenaline to brefeldin A-treated cells did not elicit a discernible change in ISC. Data obtained after permeabilisation showed that this drug causes a fall in GNa and blocks the isoprenaline-evoked increase in conductance (Fig. 1). Isoprenaline thus causes a brefeldin-A-sensitive rise in GNa in FDLE cells (Ito et al. 1997; Collett et al. 2002).
The authors thank The Wellcome TRust and Tenovus (Scotland) for their financial support.
All procedures accord with current UK legislation.