The visual system analyses the motion of 2-D patterns in two stages (Adelson & Movshon, 1982). First, orientation-selective neurons in primary visual cortex (V1) extract the motion of oriented components. Second, oriented motion signals from V1 are synthesized by neurons in visual area MT to compute speed and direction of motion (Movshon et al. 1985).
Here we examine the orientation selectivity of direction-selective and bidirectional neurons recorded in V1 of marmosets anaesthetized during preparation with saffan (I.V.) and anaesthetised and neuromuscularly blocked during recording with a respiratory mixture of 2:1 nitrous oxide and oxygen with supplementary fluothane (0-0.5 %) as required to maintain a stable ECG and synchronized EEG and intravenous delivery of 20 µg kg-1 h-1 fentanyl citrate and 100 µg kg-1 h-1 I.V. pancuronium bromide in glucose saline I.V. Procedures were licensed under the UK Animals (Scientific Procedures Act) 1986 (Derrington et al. 2002).
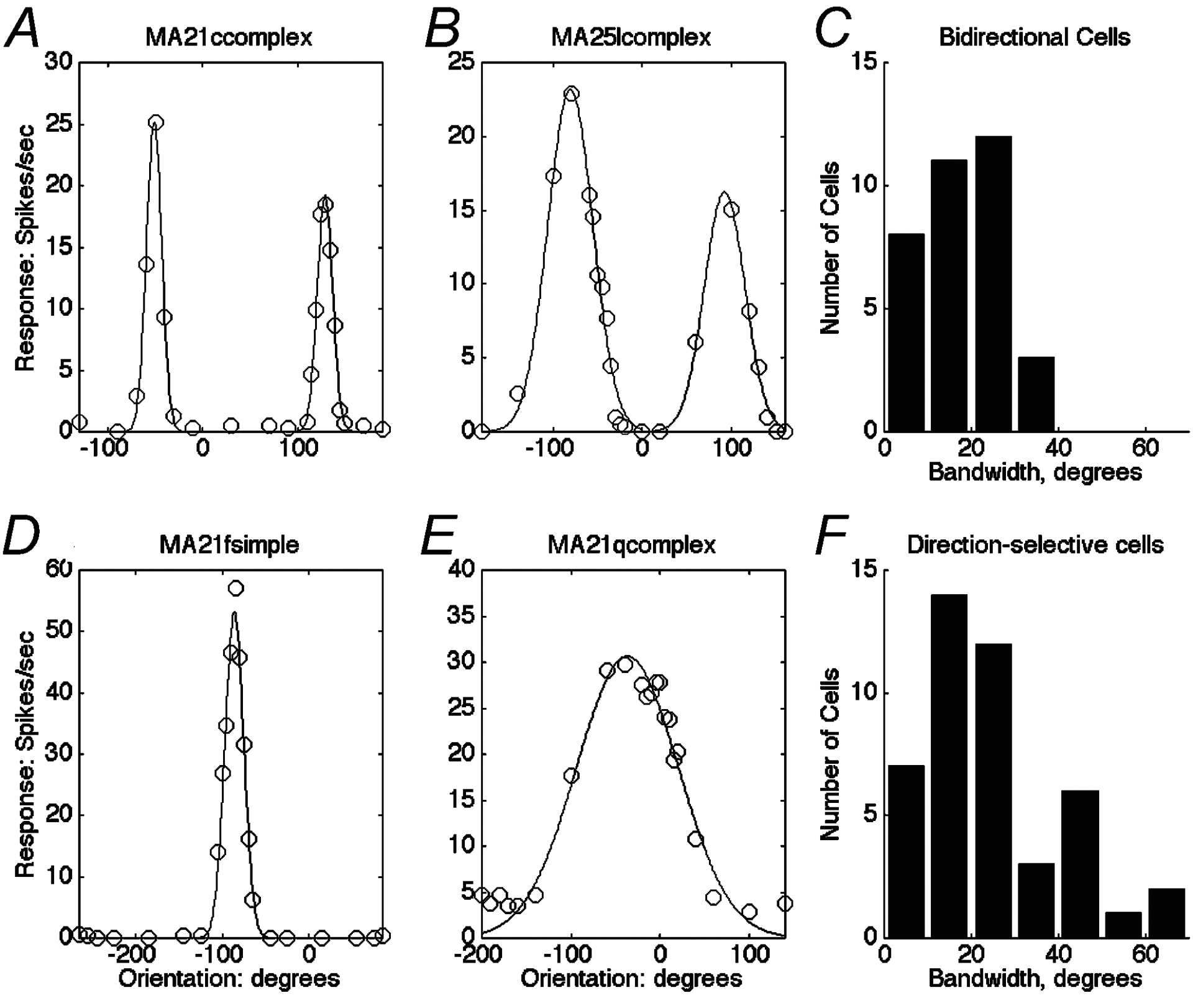
Responses to moving sinusoidal gratings depend strongly on orientation. The range of orientation bandwidths is similar to that found in macaque (Ringach et al. 2002) but the best marmoset neurons have narrower bandwidths than macaque neurons. About 25 % of direction-selective neurons have bandwidths greater than 45 deg. When tested with 2-D patterns made by adding differently oriented component gratings, broadly oriented neurons signal the motion of local features rather than the motion of component gratings.
This work was supported by The BBSRC and the Wellcome Trust.