
Innocuous warming in mice is detected by the ion channel TRPV3, expressed in the skin but not in sensory neurones (Moqrich et al. 2005). This finding may be of limited relevance to human warm sensing: a substantial number of human dorsal root ganglion (DRG) neurones express TRPV3 (Smith et al. 2002), and lack of TRPV3 expression in mouse DRG may simply reflect the fact that warm receptors are very scarce in rodents. Hatchling chicks may be a better model for studying cutaneous warm sensing: they thermoregulate only behaviourally, being unable to generate enough body heat to survive, and require high ambient temperatures (~37°C) for survival. My aim here was to investigate innocuous warm transduction in chick sensory neurones. Chicks 3 – 6 days after hatching were humanely killed, and DRG cultures and measurements of [Ca2+]i were made as described (Reid et al. 2002). Heat ramps (29 – 38°C or 29 – 53°C, 0.5°C/s) were applied with a Peltier-based system (Reid et al. 2001). Of 1321 chick DRG neurones, 100 (7.6%) responded to warming to 38°C with an increase in [Ca2+]i (ΔF/F0 0.255 ± 0.136, mean ± SD, n = 100). Thresholds were normally distributed (33.7 ± 2.0°C, n = 100), and heating to 53°C revealed a bimodal distribution of thresholds with distinct groups of neurones responding to warmth and to noxious heat (Fig. 1). Warm-sensitive neurones were significantly smaller than noxious heat-sensitive neurones (20.7 ± 3.5 μm, n = 92, vs. 24.5 ± 4.0 μm, n = 157; p < 0.001). Responses to warming depended on extracellular Ca2+ (n = 7) and were strongly inhibited by the TRPV antagonist ruthenium red (75 ± 18% inhibition at 10 μM, n = 18). Warm responses were unaffected by reducing pH to 6.5 (n = 19), suggesting TRPV1 is probably not the transducer. All warm-sensitive neurones were activated or sensitised by 50 μM 2-aminoethoxydiphenyl borane (2-APB), an agonist of TRPV1, 2 and 3 but not TRPV4 (n = 16), and by 2 mM camphor, a TRPV3 agonist which is without effect on other TRPV channels (n = 16; Fig. 2). RT-PCR showed TRPV3 mRNA to be highly expressed in chick DRG. In conclusion, chicks have substantial numbers of warm-sensitive DRG neurones, and TRPV3 plays an important role in warm transduction in these neurones. This is the first report of innocuous warm transduction in DRG neurones, and of the function of TRPV3 in the DRG. Chicks are likely to be a more useful model than rodents in understanding the roles of TRPV3 in human sensory neurones.
University of Bristol (2005) J Physiol 567P, C42
Oral Communications: Innocuous warm transduction in chick primary sensory neurones depends on TRPV3
Reid, Gordon;
1. Faculty of Biology, University of Bucharest, Bucharest, Romania.
View other abstracts by:
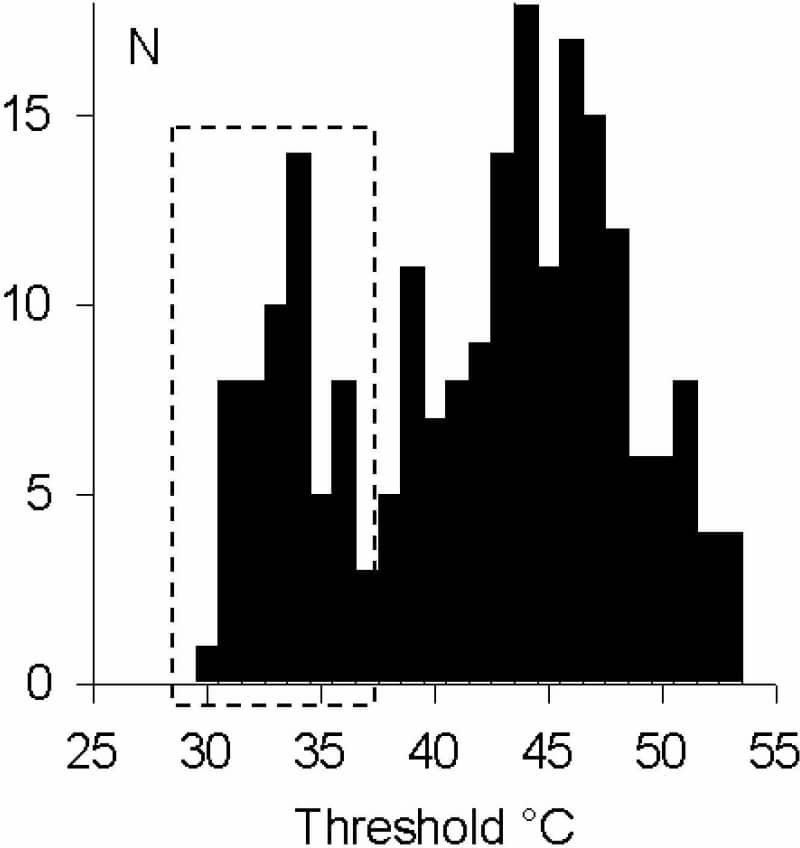
Figure 1. Thresholds of chick DRG neurones during 53°C ramps show distinct groups activated by warming (box) or by noxious heat.
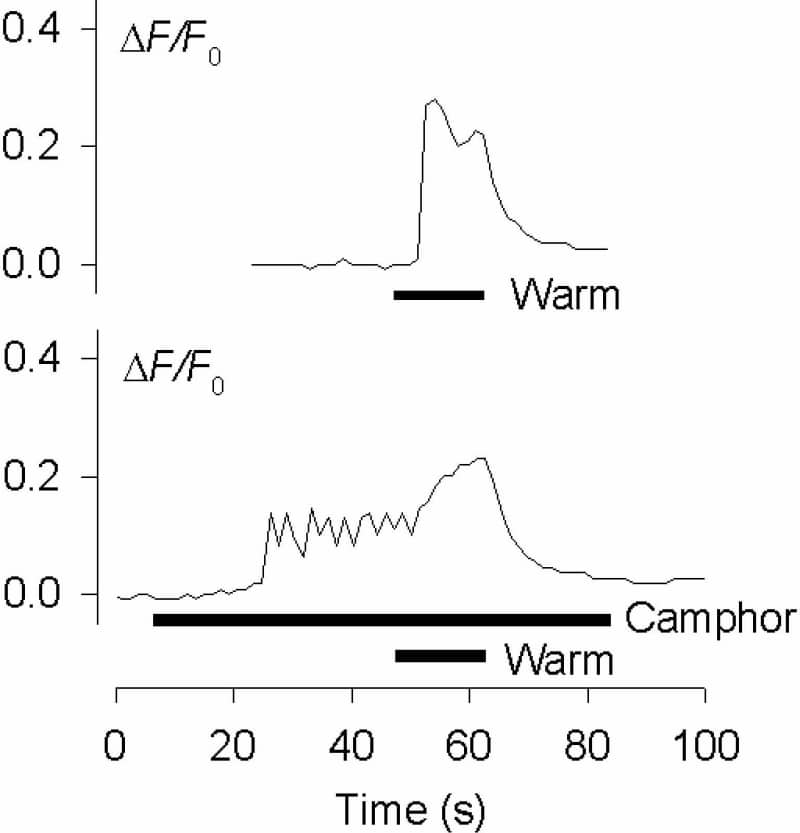
Figure 2. Activation of a chick DRG neurone by 38°C ramp (top) and by 2 mM camphor (bottom same neurone).
Where applicable, experiments conform with Society ethical requirements.