
Physiology News Magazine

Neuromuscular interaction during human walking: how do changes in muscle–tendon mechanics affect the motor control of walking?
Prolonged walking can increase the compliance of soleus tendinous tissues, which could ultimately impair the ability to recover from a sudden balance disturbance
Features
Neuromuscular interaction during human walking: how do changes in muscle–tendon mechanics affect the motor control of walking?
Prolonged walking can increase the compliance of soleus tendinous tissues, which could ultimately impair the ability to recover from a sudden balance disturbance
Features
Neil J. Cronin (1,2)
1: Center for Sensory-Motor Interaction, Department of Health Science and Technology, Aalborg University, Denmark
2: Neuromuscular Research Centre, Department of Biology of Physical Activity, University of Jyväskylä, Finland
https://doi.org/10.36866/pn.77.32

When humans walk, muscle–tendon units in the legs are constantly subjected to length changes, which are distributed between the muscle and tendinous tissues within the muscle–tendon unit. These length changes can affect the activity of afferent receptors located in the muscle. One situation where this is particularly important is when encountering an unexpected balance disturbance during walking, such as a trip or stumble. In order to avoid falling, the central nervous system must be able to detect the disturbance and initiate a response, all in a matter of milliseconds.
Investigating neural and mechanical behaviour simultaneously
A wealth of data have been presented investigating sensory feedback in humans during walking, and this has without question improved our understanding of human motor control. However, since many of the receptors that contribute to afferent feedback during walking are located in the muscle and tendinous tissues, it has become increasingly desirable to investigate the behaviour of these tissues, and to relate it to sensory feedback. One method that has facilitated experiments in this field is ultrasonography. Although this method is traditionally associated with fetal scanning, the last two decades have seen an adaptation to the methodology that has allowed the study of muscle and tendon length changes during dynamic movements. Although the method is naturally limited by the fact that it is 2-dimensional, it has nonetheless provided valuable information about muscle and tendon mechanics during movement.
Neuromuscular interaction during walking: what happens when we walk for a long time?
It has previously been shown that after a 1 h period of repeated passive stretching of the human ankle extensor muscles, the compliance of the tendinous tissues (outer tendon and aponeuroses) increases (Avela et al. 2004). If the same phenomenon were present during walking, increased tendinous tissue compliance could affect the firing behaviour of sensory receptors within the muscle–tendon unit, such as the muscle spindles. These receptors are thought to make an important contribution to muscle activation during normal walking, as well as in response to an unexpected balance disturbance (Sinkjaer et al. 2000). Therefore, changes in tendinous tissue compliance could affect the motor control of human walking.
We recently sought to examine whether changes in tendinous tissue compliance were indeed evident during human walking in a group of 11 young, healthy adults (Cronin et al. 2009). Subjects walked on a treadmill for 75 min with a portable robotic actuator attached to the ankle joint. This device is capable of eliciting rapid ankle dorsiflexion perturbations during the stance phase of walking (Fig. 1). Full details of the device are presented elsewhere (Andersen & Sinkjaer, 2003). By combining this method with ultrasonography and surface electromyography (EMG), it was possible to examine changes in both muscle fascicle length and muscle activation in the soleus muscle throughout an exercise protocol.
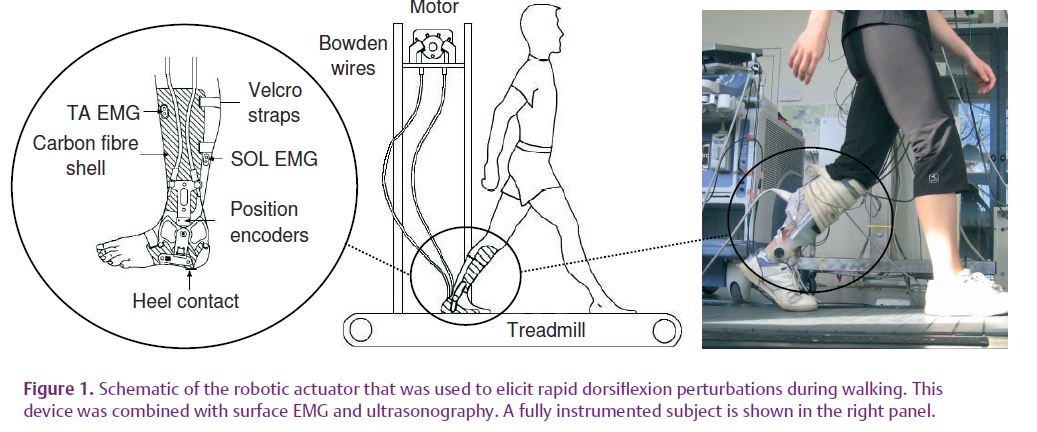
Rapid stretches were imposed at the ankle joint at three time intervals representing the early, mid and late phases of the 75 min walking protocol (Pre, Mid and Post, respectively). Between the Pre and Post intervals, the amplitude and velocity of fascicle stretch both decreased considerably (by 46% and 59%, respectively; mean values across all subjects), as did short and medium latency stretch reflex amplitudes (by 33 and 25%, respectively) in response to a constant external perturbation.
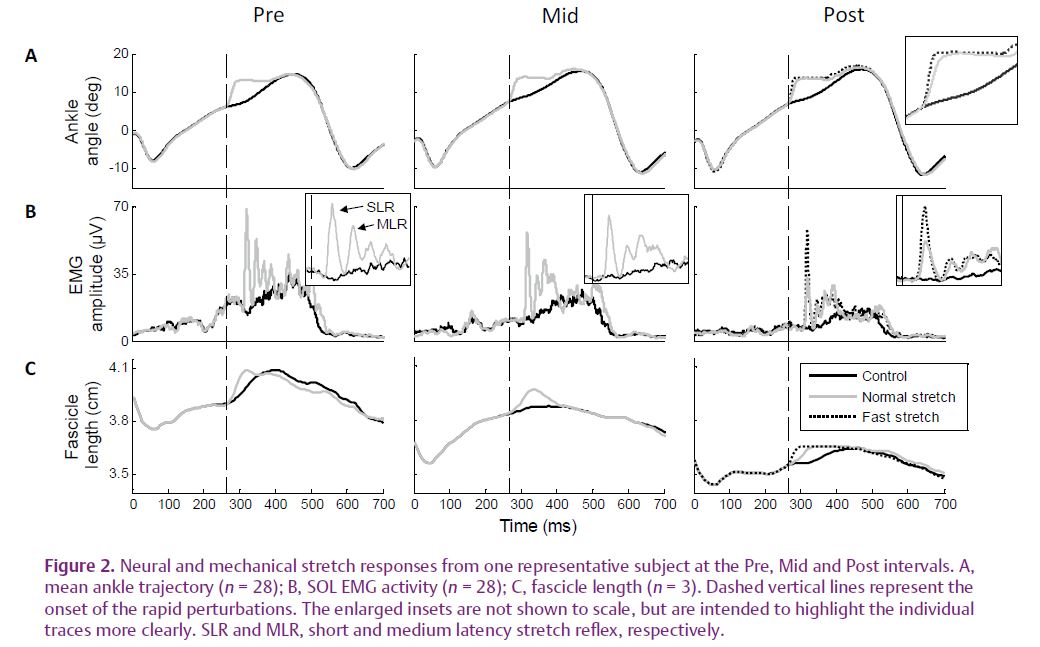
Furthermore, in response to a faster perturbation elicited at the Post interval, the amplitude of the short latency component and the velocity of muscle fascicle stretch both recovered to pre-exercise values (Fig. 2). These findings clearly show that when perturbations were elicited at the Post interval, the stretch was not transmitted as effectively to the muscle fascicles, and thus to the muscle spindles.
This would explain why short and medium latency stretch reflex amplitudes decreased, as they are at least partly mediated by type Ia and type II afferents, respectively, both of which originate in the muscle spindles. At the Post interval, we also observed a decrease in fascicle length at the point of ground contact and immediately prior to the onset of the perturbation, as well as a smaller fascicle length change throughout the stance phase. Concurrently, the range of joint motion at the ankle during walking was unchanged throughout the study. Taken together, these findings provided strong evidence for an increase in the compliance of tendinous tissues after walking in the human soleus muscle–tendon unit.
Interestingly, during normal unperturbed walking, a general decrease in ongoing soleus muscle activity was observed (‘Control’ traces in Fig. 2). Thismay be an indication that the increased tendinous tissue compliance led to a decrease in the ongoing sensory feedback from muscle receptors, which is known to contribute to muscle activity during unrestrained walking (Sinkjaer et al. 2000). As with the responses to perturbations, this is probably due to the fact that the stretch stimulus reaching the muscle fascicles (and receptors) decreased after the walking protocol.
What are the functional implications?
Based on data obtained with ultrasonography, it is known that when a muscle–tendon unit changes length, the actual length changes of the muscle and tendon components may be uncoupled (e.g. Ishikawa et al. 2005). In this context, the increase in tendinous tissue compliance that we have observed could enable the muscle fibres to contract relatively slowly (Lichtwark & Wilson, 2008), whilst the larger tendinous tissue stretch during the stance phase would facilitate the storage of elastic energy in the tendinous tissues. This could potentially increase the efficiency of the soleus muscle–tendon unit.
However, this raises an interesting series of questions. What happens in the other muscle–tendon units of the lower limb? If muscle activation decreases in soleus, how does the central nervous system compensate in order to maintain the same movement pattern? Furthermore, if the amplitude of stretch reflexes decrease after a sustained period of walking, and these responses are important for recovery from unexpected balance disturbances (Sinkjaer et al. 2000), are the responses to such disturbances compromised after prolonged walking? As the soleus muscle is just one of the plantarflexors, it is possible that activation of other synergistic muscles increases in response to decreased soleus activation. Indeed, this kind of neural compensation has been reported after higher intensity fatiguing exercise (Akima et al. 2002). This is a matter of ongoing study in our laboratory, and the results will help us to understand how the central nervous system adapts to time-dependent changes in muscle–tendon mechanics.
References
Akima H, Foley JM, Prior BM, Dudley GA & Meyer RA (2002). Vastus lateralis fatigue alters recruitment of musculus quadriceps femoris in humans. J Appl Physiol 92, 679–684.
Andersen JB & Sinkjaer T (2003). Mobile ankle and knee perturbator. IEEE Trans Biomed Eng 50, 1208–1211.
Avela J, Finni T, Liikavainio T, Niemela E & Komi PV (2004). Neural and mechanical responses of the triceps surae muscle group after 1 h of repeated fast passive stretches. J Appl Physiol 96, 2325–2332.
Cronin NJ, Ishikawa M, Af Klint R, Komi PV, Avela J, Sinkjaer T & Voigt M (2009). Effects of prolonged walking on neural and mechanical components of stretch responses in the human soleus muscle. J Physiol 587, 4339–4347. http://jp.physoc.org/content/587/17/4339. long
Ishikawa M, Komi PV, Grey MJ, Lepola V & Bruggemann GP (2005). Muscle-tendon interaction and elastic energy usage in human walking. J Appl Physiol 99, 603–608.
Lichtwark GA & Wilson AM (2008). Optimal muscle fascicle length and tendon stiffness for maximising gastrocnemius efficiency during human walking and running. J Theor Biol 252, 662–673.
Sinkjaer T, Andersen JB, Ladouceur M, Christensen LO & Nielsen JB (2000). Major role for sensory feedback in soleus EMG activity in the stance phase of walking in man. J Physiol 523, 817–827.