
Physiology News Magazine

Eccentric exercise provides new insight into sensorimotor control of muscle
Uwe Proske answers some questions about a form of exercise that leaves us stiff and sore next day, and how a study of such exercise can be used to better understand muscle and its neural control
Features
Eccentric exercise provides new insight into sensorimotor control of muscle
Uwe Proske answers some questions about a form of exercise that leaves us stiff and sore next day, and how a study of such exercise can be used to better understand muscle and its neural control
Features
Uwe Proske
Department of Physiology, Monash University, Clayton, Victoria, Australia
https://doi.org/10.36866/pn.60.14

In recent years there has been an upsurge of interest in the physiology of exercise. One reason for this is that exercise has become a part of most people’s lives as we try to maintain our fitness in the face of largely sedentary lifestyles. Then there is the rise in spectator sports, such as football, where we participate by yelling at the TV screen. With it comes a never ending search for better performances by the elite athletes shown on our screens, who are paid astronomic salaries, and whose only concern is how best to avoid exercise-induced injury. That, in turn, raises interest in the physiology of exercise.
Changes in muscle mechanical properties after eccentric exercise
In a simple classification we might identify three different kinds of exercise. The first is concentric exercise, where the contracting muscle shortens, as occurs during most movements. In isometric exercise, muscle length does not change; for example, contracting our arm muscles while grasping a support to hold ourselves upright. Finally, in eccentric exercise, where the contracting muscle lengthens, we use our muscles to control a movement, such as closing a door, without banging it, in the face of a stiff breeze. Other examples of eccentric biased activities include horse riding, downhill walking and kicking a ball. The curious thing about eccentric exercise is that it is the only form of exercise that leaves us stiff and sore the next day. Intense concentric or isometric exercise may reach the point of being painful, but there are no aftereffects a day later.
Eccentric exercise, in someone unaccustomed to it, leads to muscle damage. That, in turn, triggers an inflammatory response and the products of the inflammation are thought to sensitise muscle nociceptors, hence the pain. The primary event in the damage process is believed to be disruption of sarcomeres in the exercised muscle fibres (Morgan, 1990; Proske & Morgan, 2001). As the disruption spreads, membranes become torn leading to the uncontrolled release of calcium into the sarcoplasm. The calcium triggers a contracture in parts of the fibre. The contracture persists for several hours, presumably for as long as energy stores allow. The presence of injury contractures in damaged muscle fibres produces a rise in whole muscle passive tension (Fig. 1), which is what we experience as a sensation of stiffness.
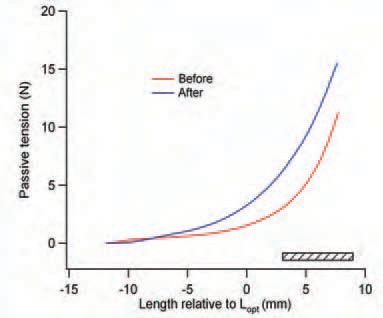
It is possible to collapse most of the extra, injury-related, passive tension by subjecting the muscle to a large passive stretch (Whitehead et al. 2003). We are not sure why. One possibility is that the stretch breaks up the region of injury contracture into smaller segments separated by parts of the fibre devoid of contractile material (Whitehead et al. 2003). It occurred to us that one possible role of the warmup stretches routinely used by athletes before exercise is to keep passive tension levels low and so help maintain a larger range of motion about the joint (Reisman et al. 2005).
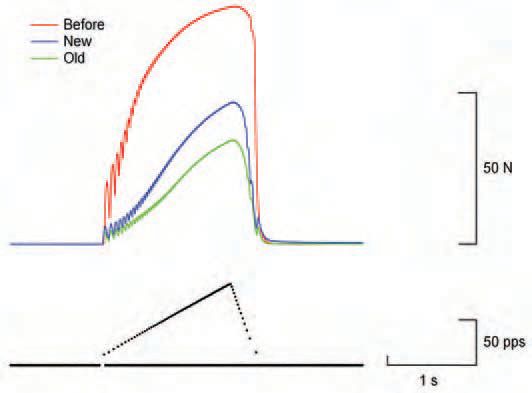
The damage to membranes impairs function of the excitation-contraction coupling system. T-tubules may shear off. A consequence for the muscle is a change in the force-frequency relation, and higher rates of stimulation are now required to achieve peak tension (Fig. 2).
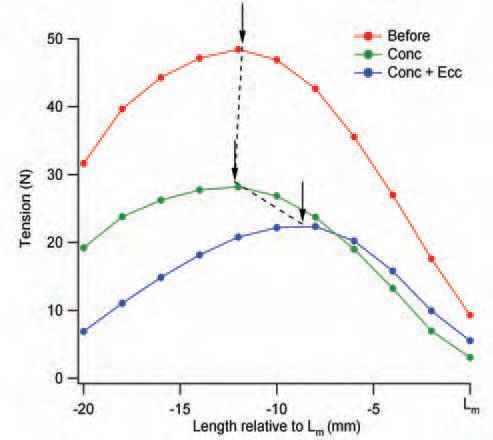
Another immediate consequence of muscle damage from eccentric exercise is a shift of the muscle’s optimum length for peak active tension, in the direction of longer muscle lengths (Fig. 3). That is, the muscle has to be stretched out further before peak tension levels are reached. The reason is that the disrupted sarcomeres are scattered at random along muscle fibres and typically lie adjacent to still functioning parts of the fibre. This means that, when the functioning sarcomeres develop force and shorten, they not only have to raise tension in the tendon, but in the adjacent non-functioning sarcomeres as well. The extra compliance represented by disrupted sarcomeres leads to a shift in the length-tension curve of the muscle in the direction of longer lengths. The shifted curve is also a little narrower, as it represents activity of a smaller number of sarcomeres in series.
It is a common misconception that muscle damage from eccentric exercise is the result of the high level of tension generated during the active stretches. It is true that during the stretch force rises above isometric levels, but it is a relatively simple matter to show that high force levels, as such, are not responsible for the damage. We have done this in two ways (Morgan et al. 2004).
First, in an animal preparation we compared the amount of muscle damage from 50 eccentric contractions in fresh unfatigued muscle with previously fatigued muscle, by determining the fall in peak active tension and the shift in optimum length.
We fatigued one muscle portion, reducing its tension by 40%, by means of a series of concentric contractions. It is known that concentric contractions are not accompanied by muscle damage. Consistent with that view was the finding that the concentric contractions were not accompanied by a shift in optimum (Fig. 3). The eccentric contractions were given to both preparations, the fatigued muscle and the fresh, unstimulated muscle. Both showed a subsequent fall in force. The size of the fall was less in the previously stimulated muscle, since many of its muscle fibres were already fatigued. More importantly, both preparations showed a similar shift in optimum length, suggestive of similar amounts of damage (Morgan et al. 2004). That raises the more general point that a drop in force, as such, is not a good indicator of how much damage has been produced, because of the additional effects of fatigue. The shift in optimum is more reliable.
In another experiment we divided the muscle nerve supply into different sized portions, in terms of the tension they generated during stimulation. Eccentric contractions of each portion led to a shift in optimum length and a fall in force. The sizes of the shifts and the relative force drops were not correlated with the amount of force generated by each piece of muscle before the eccentric contractions.
Our conclusion is that force levels are not a major determinant of the amount of damage from eccentric contractions. The factor we consider the most important is the length range over which the active stretches are carried out. If the stretch includes a portion of the descending limb of the lengthtension curve, the region of sarcomere instability (Morgan, 1990), damage is more likely.
Implications for proprioception
It is a common experience after a period of intense exercise to feel unsteady on the feet and have difficulty in carrying out skilled movements. Think of the marathon runner staggering off the track at the end of the course. This is not just a matter of muscle weakness from fatigue; there is evidence that there is a disturbance to the sense of tension and the sense of limb position after a period of eccentric exercise (Brockett et al. 1997). So the staggering athlete may be unsure of where their feet are if they are not looking at them.
We considered the possibility that eccentric exercise not only damaged muscle fibres but also disturbed the function of muscle sense organs, the muscle spindles and tendon organs. We therefore studied, in an animal model, the response properties of spindles and tendon organs after a severe bout of eccentric exercise (Gregory et al. 2002; 2004). It turned out that there was no evidence of disturbance to normal function of muscle receptors. This result meant that we had to reassess the claims about eccentric exercise and proprioception.
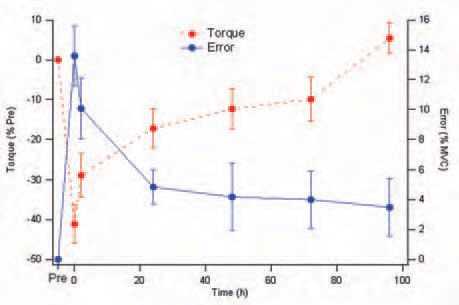
We therefore repeated a series of psychophysical experiments on human subjects and showed, specifically, that after a period of eccentric exercise subjects made large errors in an isometric force matching task (Weerakkody et al. 2003) and in a limb position matching task (Walsh et al. 2004). The size of the errors was correlated with the size of the force drop after the exercise (Fig. 4). This led us to conclude that in determining a given level of force in their muscles, or the position in space of their unsupported limbs, subjects were not just relying on information coming from sense organs in the muscle, but were using as a force or position cue the amount of effort required to carry out the task.

It is believed that the sense of effort or of heaviness is generated within the brain and, as such, is independent of feedback from the body periphery. A way to think of it is to imagine that, every time we carry out a contraction, a copy of the motor command to the muscles is sent back to sensory areas in the brain to generate a sense of effort.
During intense exercise of all kinds the muscle will fatigue, and now more effort will be required to generate a given level of muscle force. That, in turn, leads to a disturbance of the sense of force and the sense of limb position, hence the staggering marathon runner.
Concluding comments
Eccentric exercise has turned out to be an interesting topic of study. It has taught us something about the damage process in muscle following such exercise and it has led us to reassess current views about limb proprioception. There is the promise of more in the future. How is the soreness from exercise generated? Can it be minimised by training? Are muscle strains related to the damage from eccentric exercise? How can they be avoided? It is an old observation that, when one question leads to many others, we know that we are breaking important new ground.
References
Brockett C. Warren N, Gregory JE, Morgan DL & Proske U (1997). comparison of the effects of concentric versus eccentric exercise on force and position sense at the human elbow joint. Brain Res 771, 251-258.
Gregory JE, Brockett CL, Morgan DL, Whitehead NP, Proske U (2002). Effect of eccentric muscle contractions on Golgi tendon organ responses to passive and active tension in the cat. J Physiol 538, 209-218.
Gregory JE, Morgan DL, Proske U (2004). Responses of muscle spindles following a series of eccentric contractions. Exp Brain Res 157, 234-240.
Morgan DL (1990). New insights into the behaviour of muscle during active lengthening. Biophys J 57, 209-221.
Morgan DL, Gregory JE & Proske U (2004). The influence of fatigue on damage from eccentric contractions in the gastrocnemius muscle of the cat. J Physiol 561, 841-850.
Parikh S, Morgan DL, Gregory JE and Proske U (2004). Lowfrequency depression of tension in the cat gastrocnemius muscle after eccentric exercise. J Appl Physiol 97.
Proske U and Morgan DL (2001). Muscle damage from eccentric exercise: Mechanism, mechanical signs, adaptation and clinical applications. J Physiol 537, 333-345.
Reisman S, Walsh LD & Proske U (2005). Warm-up stretches reduce sensations of stiffness and soreness after eccentric exercise. Med Sci Sports Exerc 37, 929-936
Walsh LD, Hesse CW, Morgan DL, Proske U (2004). Human forearm position sense after fatigue of elbow flexor muscles. J Physiol 558, 705-715.
Weerakkody NS, Percival P, Morgan DL, Gregory JE, Proske U (2003). Matching different levels of isometric torque in elbow flexor muscles after eccentric exercise. Exp Brain Res 149, 141-150.
Whitehead NP, Morgan DL, Gregory JE and Proske U (2003). Rises in whole muscle passive tension of mammalian muscle after eccentric contractions at different muscle lengths. J Appl Physiol 95, 1224-1234.