The myocardial force-frequency relation (FFR) is positive at steady-state in mammals except the rat where it is negative (Layland & Kentish, 1999). In frogs, FFR is positive up to a certain optimum frequency. We propose a mechanism involving a single protein to explain the FFR in the frog-ventricle.
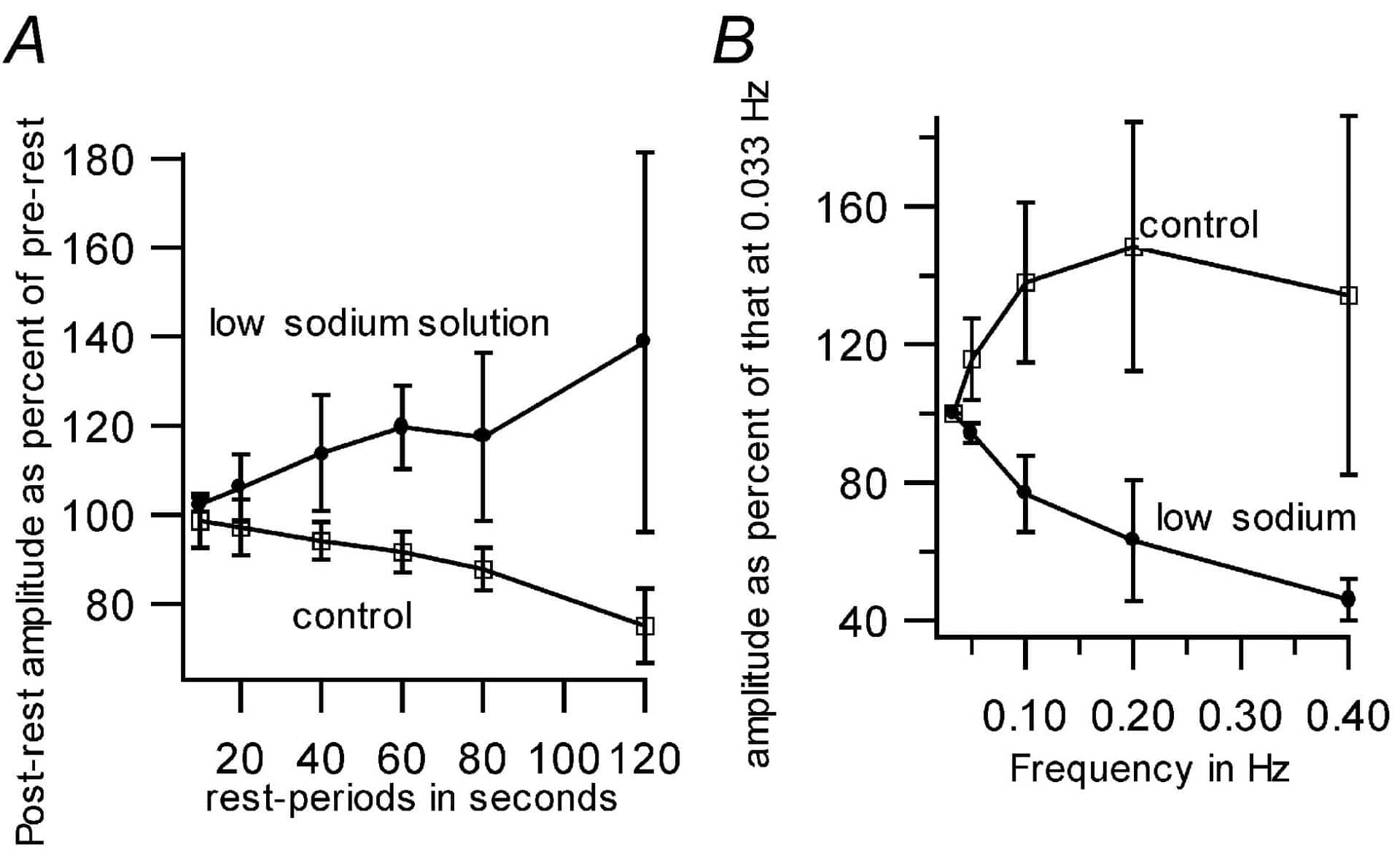
Circular ventricular strips, made from isolated hearts of frogs (Rana hexadactyla) anaesthetised with ether and then pithed, were used. In one set of experiments, the strips were paced at 0.2 Hz and the effect of rest on post-rest beat amplitude was studied. Under control conditions, there was rest-induced decay (RID) of amplitude. This is due to a steady diastolic leak of calcium from sarcoplasmic reticulum (SR) to the outside (Subramani et al. 2002). We report here that RID was not affected by the following interventions: adrenaline (10 µM; augments L-type calcium currents and SR calcium pump) ryanodine (10 µM; ryanodine receptor blocker), neomycin (10 µM; inhibitor of IP3 formation) lanthanum (10 µM; sarcolemmal calcium-pump blocker), and nickel (40 µM; T-type calcium channel blocker). The proteins whose functions are modified by these agents are therefore not involved in the RID phenomenon. In another protocol, the frequency was stepped up from 0.03 Hz to about 0.5 Hz and then stepped down. The force at a particular frequency was averaged from the two cycles. There was a positive FFR under control conditions. The interventions mentioned above which did not change the RID phenomenon did not affect the positive FFR either. When the direction of the sodium-calcium exchanger (NCX) which is normally calcium-extrusive in diastole (Bogdanov et al. 2001) was reversed with (a) low sodium concentration (40 mM, osmolarity corrected with sucrose), (b) high calcium concentration (5 mM) and (c) ouabain (10 µM), the post-rest beat was larger than the pre-rest (rest-induced potentiation, RIP). In addition, the FFR turned negative. With reversal, NCX becomes calcium-acquisitive in diastole. The rest periods now help in accumulating calcium into SR and hence the RIPs.
To summarize, when there is RID, FFR is positive, and whenever there is RIP, FFR turns negative (Fig. 1). We hypothesize that when the conditions favour a net leak of calcium from SR during diastole (precisely when the conditions favour the calcium-extrusive mode of NCX), FFR is positive. An increase in frequency shortens diastole and therefore the diastolic calcium leak, thereby augmenting force. On the other hand, interventions which lock the NCX in reverse mode, causing a net uptake of calcium in diastole, were able to change the pattern of RID to RIP and converted the FFR from positive to negative. With net diastolic calcium uptake, lower frequencies can help increase force. In conclusion, the force-frequency relation is dependent on the direction of the sodium-calcium exchanger.
S. Subramani thanks the Wellcome trust for the travel grant.