
Many neurons in sensory areas of the brain are directionally selective. In visual cortex, cells commonly fire more in response to oriented light or dark bars sweeping in one direction compared with the reverse[1]. Analogous directionality is seen in somatosensory[2] and auditory cortex. The mechanisms are still debated. In retina and some invertebrate systems, local dendritic computations support directionality: could this play a role in mammalian neocortex? Most inputs into cortical neurons arrive on sub-micron diameter “thin” dendrites, such as basal & apical oblique dendrites, which can also fire NMDA spikes in response to sufficient excitatory synaptic stimulation[3]. NMDA spikes are ideally suited to the computation of directionality: a) Focally-evoked NMDA spikes in basal & oblique dendrites exhibit pronounced spatial gradients in both amplitude & glutamate threshold[4]: a proximal NMDA spike can produce a 7-fold bigger somatic voltage swing than a distal NMDA spike in the same dendrite & requires ~5 times as much glutamate to cross threshold (in large layer 5 cells). b) Depolarisation reduces the glutamate threshold for an NMDA spike: a distal NMDA spike can help a more proximal one in the same dendrite cross threshold (co-operativity)[4]. Simulations predict these features should allow single dendrites to generate bigger responses following distal-to-proximal (DP) sweeps of glutamate stimuli compared with exactly the same stimuli in proximal-to-distal (PD) order; also, strategically placed and timed inhibition should enhance this directionality. To test this, combined whole-cell recording, 2-photon Ca2+ imaging, patterned 2-photon glutamate uncaging[5] and focal GABA iontophoresis experiments were performed on layer 5 pyramidal cells in rat somatosensory cortical slices. Single basal and oblique dendrites were stimulated with sweeps of glutamate spots in both directions (≤90 x 0.5-1 ms uncaging spots near spines, sweep duration <230 ms), in ~½ of cases comparing responses with/without 5 ms GABA pulses onto the proximal part of the same dendrite. Single dendrites could exhibit up to 7-fold or more directionality (DP response/PD response); average maximum achieved directionality of each dendrite = 2.84 ± 1.73 (mean ± sd; range 1.3-8.8, n=21 dendrites with >100 μm stimulated; no action potentials). Directionality depended on sweep speed, baseline voltage & spatial pattern of stimuli, & increased with length stimulated. Directionality was enhanced by proximal inhibition before PD sweeps (IPD), but after DP sweeps (DPI). This increased: a) the range of stimuli giving directionality, b) the directionality of a given excitatory pattern (1.62±1.07-fold mean increase per pattern, n=39 patterns). Over 11 dendrites, the maximum enhancement of directionality by inhibition averaged 1.84±1.07-fold (range 1.1-4.2).
Cardiff University (2009) Proc Physiol Soc 17, C10
Oral Communications: Single dendrite directional selectivity in neocortical pyramidal neurons, assisted by NMDA spikes and excitatory-inhibitory interactions
G. Major1, K. D. Fox1
1. School of Biosciences, Cardiff University, Cardiff, Wales, United Kingdom.
View other abstracts by:
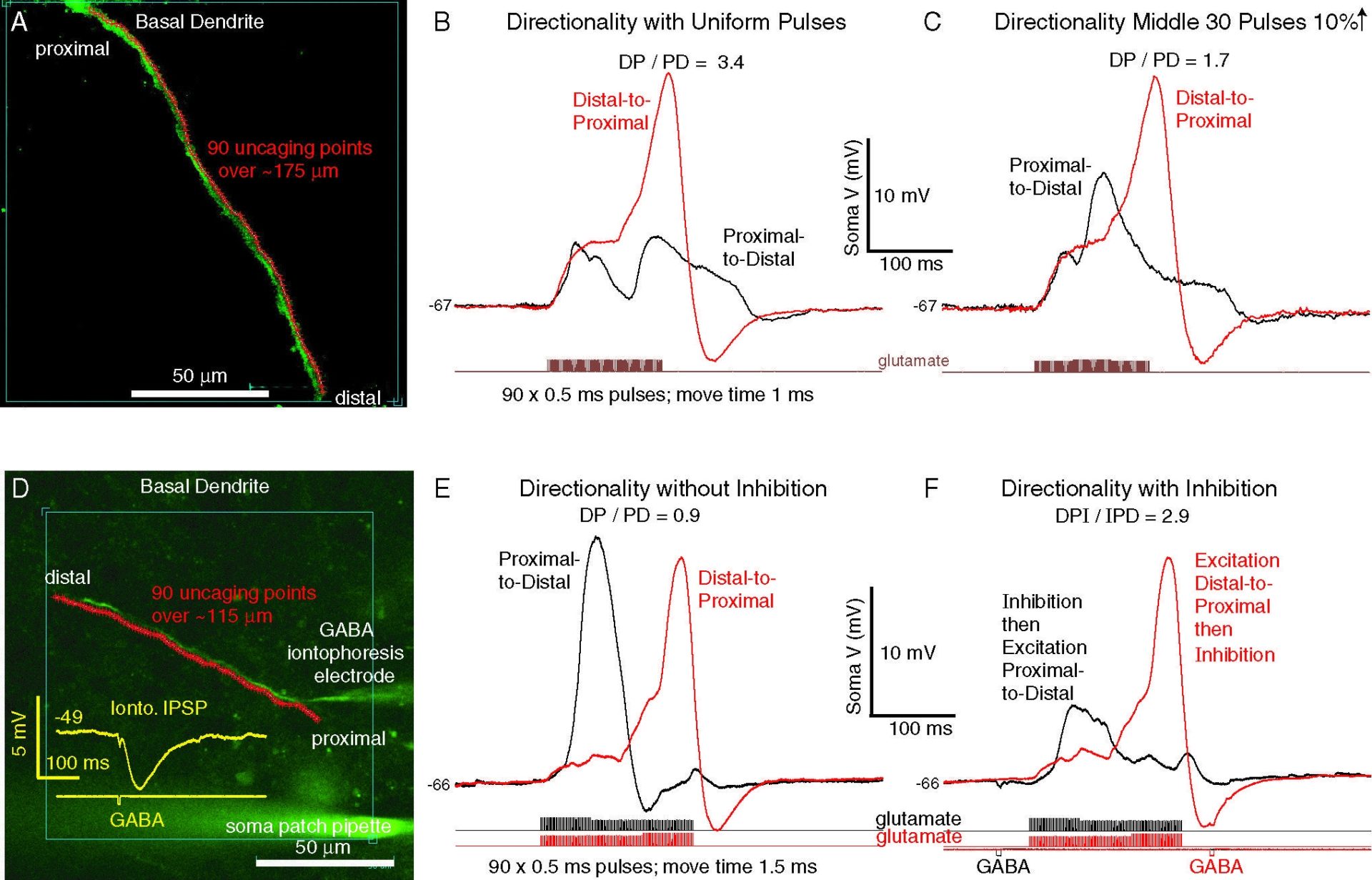
Figure 1. Examples of directionally-selective responses to patterned uncaging sweeps of glutamate spots. Top: excitation alone. Directionality sensitive to exact stimulus pattern. Bottom: timed proximal inhibition (brief iontophoretic GABA pulse) can enhance directionality.
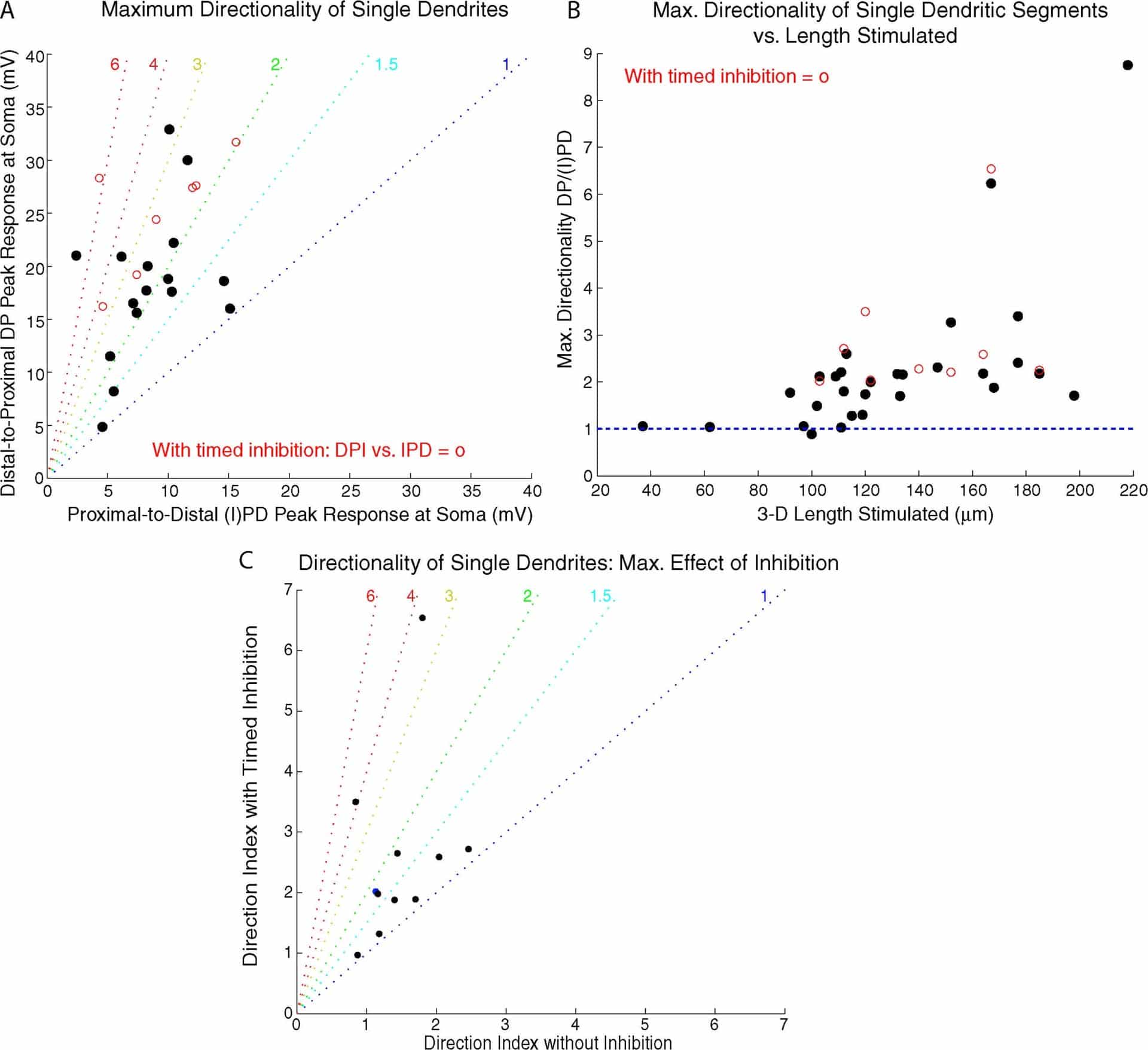
Figure 2. A. Maximum directionality of different dendrites. Red circles = cases with timed inhibition giving biggest directionality. B. Maximum directionality vs. length of segment stimulated. C. Maximum effect of timed inhibition on directionality of 11 dendrites tested with glutamate sweeps +/- GABA pulse.
Where applicable, experiments conform with Society ethical requirements.