
Physiology News Magazine
Have the carotid chemoreceptors anything to do with the control of breathing during exercise in man?
In man, breathing matches (or exceeds) metabolic rate (the rate of oxygen consumption) from rest to maximum exercise. How the brain achieves this remains one of the biggest unanswered questions in physiology.
Features
Have the carotid chemoreceptors anything to do with the control of breathing during exercise in man?
In man, breathing matches (or exceeds) metabolic rate (the rate of oxygen consumption) from rest to maximum exercise. How the brain achieves this remains one of the biggest unanswered questions in physiology.
Features
Michael Parkes
University of Birmingham, UK
https://doi.org/10.36866/pn.94.22
The traditional belief is that breathing matching metabolic rate is somehow controlled by the carotid chemoreceptors in man. Yet since their discovery around 80 years ago, experimental evidence has consistently failed to support this. The carotid chemoreceptors are in the wrong anatomical location to detect metabolic rate, receive no known signal linking them to metabolic rate and their denervation has remarkably little effect on breathing matching metabolic rate. Intriguingly in 2014 we still remain unable to establish either that they control or that they do not control breathing matching metabolic rate during exercise in man.
The carotid chemoreceptors are located bilaterally at the bifurcation of the common carotid arteries and connected to the brain via a branch of the glossopharyngeal nerve (IX). Directly stimulating them causes an immediate increase in breathing. Their principal stimuli are arterial hypoxia, hypercapnia or acidaemia. Moreover they are thought to be the only chemoreceptors in man that can detect hypoxia to stimulate breathing (because patients without them fail to increase their breathing in hypoxia). Their discovery in dogs in around 1927 was believed to be so fundamental that it won Corneille Heymans the Nobel Prize in Physiology or Medicine in 1938. It was believed to be only a matter of time before their fundamental role in controlling breathing would be established in man. Yet such experimental evidence consistently fails to materialise.
It is reasonable to assume that one continuous process explains breathing matching metabolic rate from rest to maximum metabolic rate and the easiest way to establish that breathing is matching metabolic rate is from the stability of arterial blood gas levels. To establish if the carotid chemoreceptors have any role in this matching, classical scientific method (Stein & Stoodley, 2006; Parkes, 2013) would seek to obtain positive results with the following three key experimental approaches:
- stimulation (their stimulation at rest should be able to mimic the minute ventilation levels of maximum exercise)
- ablation (their destruction should abolish matching and prevent minute ventilation ever rising appropriately during exercise)
- recording (their afferent nerve discharge should always be proportional to metabolic rate – from rest to maximum exercise)
If carotid chemoreceptors are important, the effects of stimulation and ablation might be detectable at rest, but should be most obvious at the highest metabolic rates. Moreover their role should be particularly easy to see in humans. This is because it is easy to ask humans to exercise at maximum metabolic rate and because in humans only one set of peripheral chemoreceptors – the carotid chemoreceptors – mediate the hypoxic ventilatory response. In contrast, such experiments are much more difficult in other species, where measurements at high metabolic rates are rarely attempted and the existence of multiple peripheral chemoreceptors mediating the hypoxic ventilatory response makes denervation experiments much harder to undertake and to interpret.
If the carotid chemoreceptors have no role in matching, the above experiments should show no effects (i.e. their results would be negative). Unfortunately, whereas appropriate positive results are essentially self-validating, it is much more difficult to validate negative results. Either the negative experiment was in some way inadequate (e.g. poorly controlled with insufficient numbers), or multiple control factors exist, or it is genuinely negative because the ablated factor has no role.
Since 1927, the difficulty has always been in how best to interpret the essentially negative results of the above three key experimental approaches with carotid chemoreceptors in man.
Recorded arterial blood gases fail to change appropriately during exercise

Figure 1 (Sun et al. 2001) shows no substantial rise in arterial PCO2 (PaCO2) during exercise in man. Indeed it actually falls during more severe exercise. Similarly no consistent fall in PaO2 has ever been measured at maximum exercise (and the occasional observed falls are insubstantial). So how can the carotid chemoreceptors ever ‘know’ what metabolic rate is? These facts have been known in outline since before the carotid chemoreceptors were discovered. Yet increasingly better recording experiments are still published in case something different might appear. Are these negative results definitive? Investigation of more subtle functions of PaCO2 too has failed to reveal anything useful (CO2 oscillations apparently disappear and CO2 sensitivity fails to rise sufficiently – or at all – during exercise (Parkes, 2013)). Yet some new function of arterial blood gases, or some new arterial chemoreceptor stimulant might still be found (or not, if they do not exist!).

Figure 2 shows that carotid chemoreceptors in man are actually in completely the wrong location to measure metabolic rate. It has been known since the 1920s that it is systemic venous blood gas levels that change in proportion with metabolic rate (and see Fig. 1). But systemic venous chemoreceptors have not yet been found in man and the search for them seems to have been abandoned many years ago.
Carotid chemoreceptor ‘stimulation’ in man fails to achieve the breathing of maximum exercise
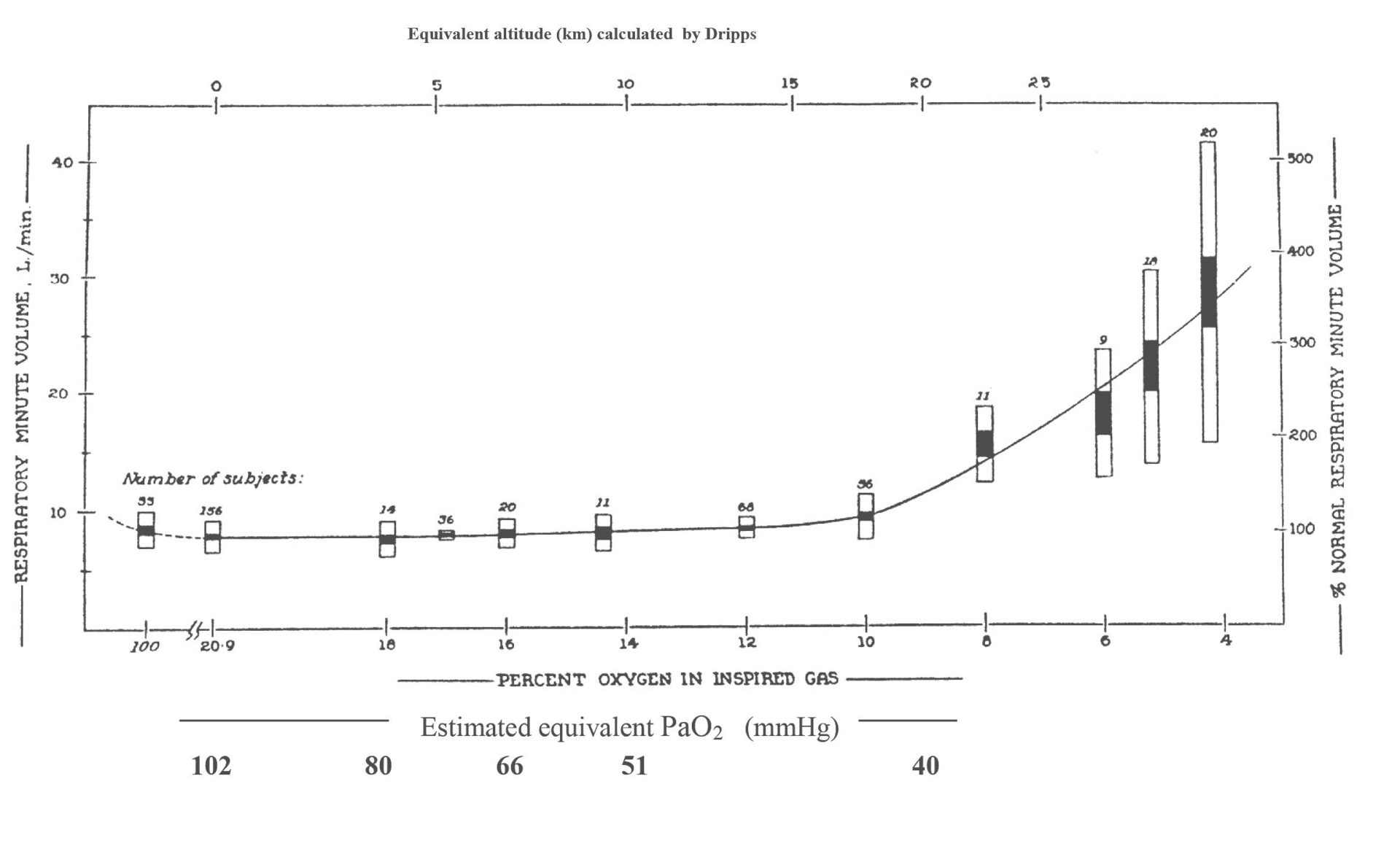
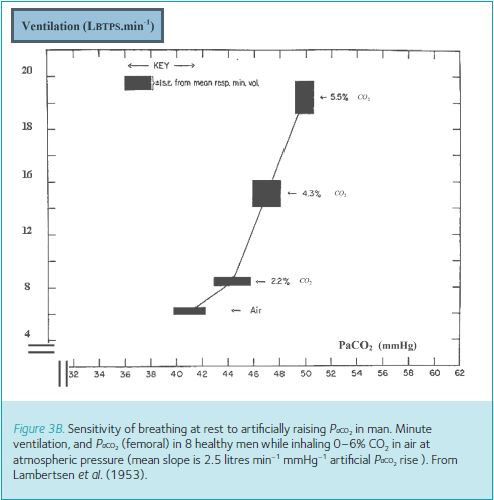
In man it remains impossible to stimulate carotid chemoreceptors directly, but hypoxia and hypercapnia are their well-known stimulants. Figure 3A and B (Dripps & Comroe, 1947; Lambertsen et al. 1953) shows how good these are at stimulating breathing to the levels seen at maximum exercise (100–150 litres (BTPS) of air per minute). Breathing ~6% oxygen achieves only ~20 litres (BTPS) of air per minute and with any more severe hypoxia, humans start to pass out. Similarly even breathing ~7% CO2 achieves only ~40 litres (BTPS) of air per minute. It can be argued that by breathing more severely hypercapnic gases, or even by combining this with hypoxia (i.e. breathing asphyxiating gas mixtures), greater breathing levels could be achieved. But this seems pointless since such asphyxiating arterial blood gas levels do not occur during exercise.
Carotid chemoreceptor ablation barely changes matching during exercise
If the carotid chemoreceptors were solely responsible for matching in man, then it might be expected that ablating them bilaterally would have major effects on blood gas levels at rest and catastrophic effects during exercise. Breathing and hence blood gases might become so unstable that other responses might be triggered (Parkes, 2013), e.g. unconsciousness at PaO2 levels below ~25 mmHg and/or PaCO2 levels above about 90 mmHg, or hypocapnic tetany at PaCO2 levels below 20 mmHg (hyperoxia being innocuous in the short term).
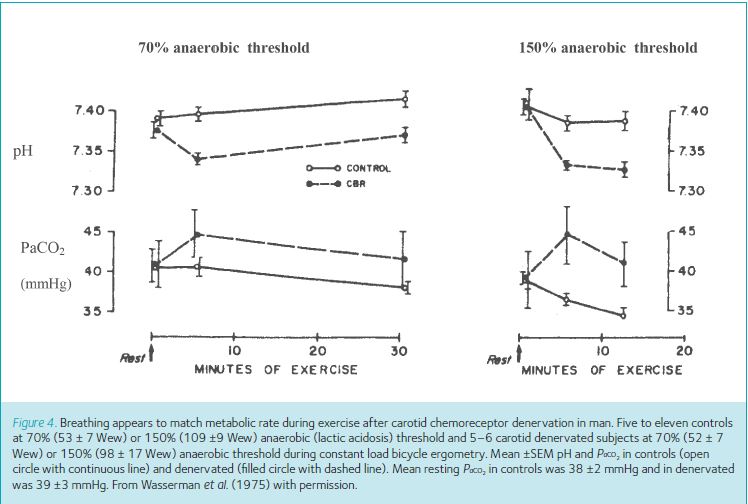
Figure 4 (Wasserman et al. 1975) shows how little the matching of breathing with metabolic rate is disturbed (i.e. how relatively stable is PaCO2) during exercise in patients with bilateral carotid chemoreceptor denervation. (Compare the PaCO2 levels in Fig. 4 with those required to stimulate breathing in Fig. 3B). Even if the data in Fig. 4 are accepted as evidence that there is a small change in PaCO2, we still do not know whether this change was caused by their asthma or by the denervation (because the controls were different subjects, not the same patients before and after denervation).
Intriguingly this negative experiment (denervation making such a small difference) has never been accepted as evidence that carotid chemoreceptors have no role. This is despite the accompanying negative evidence from the above recording and stimulation experiments. This result has always been interpreted instead as showing that carotid chemoreceptors still do control breathing during exercise, but something else too is equally important and can immediately take over! In fact we cannot yet distinguish between these two interpretations, so the correct conclusion of this negative ablation experiment so far is ‘inconclusive without further investigation’.
Conclusions
The unquestioned belief in Heymans’s carotid chemoreceptors has always trumped the experimental evidence in man that has been obvious ever since. And intriguingly, because of the difficulty in validating negative results, neither can we establish definitively that this belief is wrong!
Suppose one day we might be able to record the afferent activity from the carotid chemoreceptors in man. If it fails to change in proportion to metabolic rate, then we know finally that this belief is incorrect. If it does increase with metabolic rate then we know that all the negative stimulation and ablation results to date are false.
Has this belief really served us so well over the last 80 years that it should continue to be propagated without question in some textbooks, reviews and scientific papers? It may well have done no harm in the sense that it may never have killed a patient nor helped anyone win (or lose) a race. But might it also be preventing recognition of inconveniently novel hypotheses? Might we make more progress over the next 80 years if we started to question this belief more overtly, rather than just continuing to accept and to propagate it?