
Physiology News Magazine

HIMaging the kidney: High resolution helium ion microscopy
Various imaging techniques have been widely used to investigate kidney structure and function. Helium Ion scanning Microscopy (HIMaging) constitutes a major technological breakthrough that can potentially push the imaging resolution limit beyond that of any other currently available technique.
Features
HIMaging the kidney: High resolution helium ion microscopy
Various imaging techniques have been widely used to investigate kidney structure and function. Helium Ion scanning Microscopy (HIMaging) constitutes a major technological breakthrough that can potentially push the imaging resolution limit beyond that of any other currently available technique.
Features
Teodor G. Paunescu, Sylvie Breton & Dennis Brown
Massachusetts General Hospital and Harvard Medical School, USA
https://doi.org/10.36866/pn.95.32
Maintaining body homeostasis for pH, fluid osmolality and volume, and electrolyte balance is accomplished by the kidneys through a number of active and passive membrane transport mechanisms that occur in specialized cell types located in distinct tubule segments (Brenner & Rector, 2012). Accordingly, the intricacy of kidney structure underlines this functional complexity. For example, most of the filtered salt, amino acids and glucose are reabsorbed in the proximal convoluted tubule (PT) and thick ascending limb of Henle, whereas water is reabsorbed in the PT, thin descending limb, and principal cells of the cortical and medullary collecting duct (CD). On the other hand, acid–base homeostasis is achieved by the kidneys, in concert with the lungs, via proton secretion and bicarbonate reabsorption in the PT and by intercalated cells of the CD.
Given the strong structure–function relationship of renal epithelial cells, exploring kidney morphology, structure and ultrastructure, including cell membrane topography, has been for decades a major way of investigating their functional characteristics. For higher resolution, conventional bright-field and fluorescence microscopy gives way to scanning or transmission electron microscopy, or to atomic force microscopy.
In recent years, scanning helium ion microscopy (HIM) has emerged as a technology that can potentially offer higher resolution imaging compared to electron microscopy (Ward et al. 2007). This is achieved by scanning the sample with a narrow beam of high energy helium ions instead of electrons, thus allowing the beam to focus to subnanometer dimensions. HIM has been used significantly in materials science, but only recently has it also been applied to study biological samples, such as butterfly scales (Boden et al. 2012), fly cuticle (Boseman et al. 2013), mammalian articular cartilage (Vanden Berg-Foels et al. 2012), cancer cells (Bazou et al. 2011) and renal epithelial cells (Rice et al. 2013).
Tissue preparation for HIM
Preparation of kidney samples from adult male Sprague–Dawley rats and adult male and female C57/BL6 mice for HIM imaging has been described in detail previously (Rice et al. 2013). Briefly, it involves tissue fixation by transcardial perfusion followed by immersion in aldehyde fixatives, such as glutaraldehyde (GA), modified paraformaldehyde (PFA)–lysine–periodate (PLP) (Rice et al. 2013), or modified Karnovsky’s fixative containing 2.5% GA and 2% PFA. Small pieces of fixed tissues are then dehydrated in graded alcohol (methanol or ethanol) and subjected to critical point drying. An important feature of HIM, compared to conventional scanning electron microscopy (SEM), is that the kidney samples do not need to be covered with a conducting metal coat, thus allowing for a clearer visualization of the actual cell surface architecture. HIM was performed as previously described (Rice et al. 2013), yielding high resolution images of the renal glomerulus, PT and CD, whose quality and sharpness greatly exceed those of images obtained by standard SEM (see for example Madsen et al. 1988).
HIMaging of the glomerulus
The renal glomerulus is the site of plasma ultrafiltration across capillaries. The blood vessels in the glomerulus are surrounded by podocytes, specialized epithelial cells characterized by the presence of highly interdigitated, branching foot processes. The spaces between these processes represent the location of the filtration slit diaphragm (Brenner & Rector, 2012). Figure 1A shows the glomerular podocyte foot processes imaged by HIM in the cortex of a rat kidney. Increasing the magnification allows visualization of the podocyte filtration slit, which appears as a ladder-like structure stretching between adjacent foot processes (Fig. 1B). The blood-facing surface of endothelial cells of the renal glomerular capillary from a rat kidney (Fig. 2) shows the tight junction separating two adjacent cells and the round fenestrations that typify these glomerular endothelial cells. These fenestrations are 60–90 nm in diameter (Rice et al. 2013) and they render the endothelium freely permeable to water and small solutes.

HIMaging of renal tubules
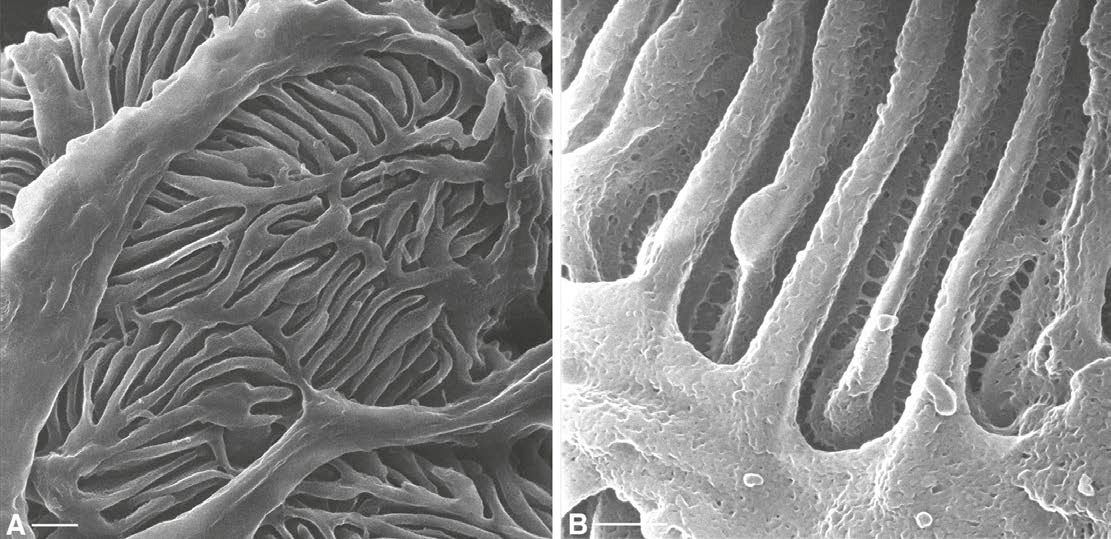
The proximal convoluted tubule (PT), the first segment of the renal tubule after the glomerulus, is highly active in membrane transport processes that undertake the reabsorption of the ultrafiltrate that is delivered to the PT from the glomerulus. The PT is involved in the reabsorption of water, ions and organic solutes, including glucose and amino acids (Brenner & Rector, 2012). PT cells are characterized by the presence of a highly developed brush border – a layer of densely packed microvilli that cover the apical surface of the cell, considerably increasing its surface area (Fig. 3A). The brush border is seen in great detail in HIM images: Fig. 3B shows the apical pole of a PT cell from a rat kidney. Increasing the magnification reveals the detailed morphology of PT brush border microvilli, including microscopic depressions on their lateral surface, which we termed micropits, and also thin filaments that appear to connect adjacent microvilli (Fig. 3C). When imaged from the luminal side, the tips of the microvilli are seen aggregating together in clusters of variable numbers (Fig. 3D).

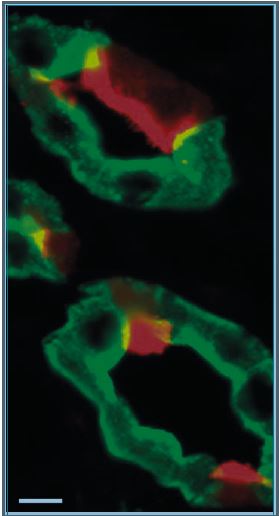
The collecting duct (CD) is the most distal portion of the renal tubule, and is active in the transport of water, sodium, chloride, potassium, protons and bicarbonate. The CD comprises two distinct cell types that fulfil two major physiological functions: principal cells (PCs) mediate water reabsorption, thus regulating body water balance, and intercalated cells (ICs) are responsible for proton secretion and bicarbonate reabsorption, and hence body pH homeostasis, as reviewed extensively elsewhere (Wagner et al. 2004; Fenton & Knepper, 2007; Brown et al. 2012). PCs and ICs express different sets of transport proteins (Fig. 4), including the proton pump V-ATPase (red) located in the apical pole of ICs, and aquaporin 2 (green) located in the apical and basolateral membrane of PCs. ICs and PCs also show considerable differences in their respective morphologies, which are highlighted very clearly by HIM. A PC is characterized by the presence of a single central cilium, on average around 3 μm long (Rice et al. 2013) and numerous short microvilli, whereas an IC has no cilium, but instead an elaborate network of apical microplicae and infoldings (Fig. 5A). The size and number of these microplicae correlate with the level of activation of transmembrane transport in IC, which in turn correlates with the rate of exocytosis of specialized vesicles that contain the V-ATPase (Wagner et al. 2004; Brown et al. 2009). In this respect, the IC shown at higher magnification in Fig. 5B is more representative of a stimulated animal –e.g. in response to elevated cAMP levels (Paunescu et al. 2010) – although different ICs usually reveal various degrees of activation even under ‘baseline’ conditions.
Given the number and size of the microplicae, the flat region of the IC apical plasma membrane is barely visible (Fig. 5C). High resolution HIM imaging of a PC (Fig. 5D) better illustrates their stubby apical microvilli and exposes subapical cytosolic structures. The depressions seen on the surface of the PC membrane, frequently found at the base of the microvilli, are thought to constitute the sites of exocytic or endocytic events.
Conclusion
We applied the relatively new technique of helium ion scanning microscopy successfully to study the rodent kidney. Our results show that this novel technology advances the field appreciably by improving the resolution with which uncoated biological specimens can be imaged. We refer to the resulting micrographs as ‘HIMages’. Given the remarkable quality, detail definition, and sharpness of such HIMages, we anticipate that expanding the use of this technology for investigating biological samples has the potential to further our understanding of their detailed structure and function.
Acknowledgements
We are grateful to Alfred N. Van Hoek (University of Utah) and William L. Rice (Massachusetts General Hospital and Harvard Medical School) for their contribution to sample preparation and HIM imaging, to Chuong Huynh, Bernhard Goetze and Larry Scipioni (Carl Zeiss Microscopy) for providing access to their helium ion microscope and for assistance with imaging, to Paul Kelly (Salem State University) and to Ann Tisdale (Schepens Eye Research Institute) for providing access to their critical point drying apparatus.

References
Bazou D, Behan G, Reid C, Boland JJ & Zhang HZ (2011). Imaging of human colon cancer cells using He-Ion scanning microscopy. J Microsc 242,
290–294.
Boden SA, Asadollahbaik A, Rutt HN & Bagnall DM (2012). Helium ion microscopy of Lepidoptera scales. Scanning 34, 107–120.
Boseman A, Nowlin K, Ashraf S, Yang J & Lajeunesse D (2013). Ultrastructural analysis of wild type and mutant Drosophila melanogaster using helium ion microscopy. Micron 51, 26–35.
Brenner BM & Rector, F (2012). The Kidney, 9th edn. Elsevier/Saunders, Philadelphia.
Brown D, Bouley R, Paunescu TG, Breton S & Lu HA (2012). New insights into the dynamic regulation of water and acid/base balance by renal epithelial cells. Am J Physiol Cell Physiol 302, C1421–C1433.
Brown D, Breton S, Ausiello DA & Marshansky V (2009). Sensing, signaling and sorting events in kidney epithelial cell physiology. Traffic 10, 275–284.
Dantzler WH (2003). Regulation of renal proximal and distal tubule transport: sodium, chloride and organic anions. Comp Biochem Physiol A Mol Integr Physiol 136, 453–478.
Fenton RA & Knepper MA (2007). Mouse models and the urinary concentrating mechanism in the new millennium. Physiol Rev 87, 1083–1112.
Madsen KM, Verlander, JW & Tisher C C (1988). Relationship between structure and function in the distal tubule and collecting duct. J Electron Microsc Tech 9, 187–208.
Paunescu TG, Ljubojevic M, Russo LM, Winter C, McLaughlin MM, Wagner CA, Breton S & Brown D (2010). cAMP stimulates apical V-ATPase accumulation, microvillar elongation, and proton extrusion in kidney collecting duct A-intercalated cells. Am J Physiol Renal Physiol 298, F643–654.
Rice WL, Van Hoek AN, Paunescu TG, Huynh C, Goetze B, Singh B, Scipioni L, Stern LA & Brown D (2013). High resolution helium ion scanning microscopy of the rat kidney. PLoS One 8, e57051.
Vanden Berg-Foels W, L Scipioni, C Huynh & X Wen (2011). High resolution visualization of the articular cartilage collagen network by helium ion microscopy. Microsc Microanal 17, 282–283.
Vanden Berg-Foels WS, Scipioni L, Huynh C & Wen X (2012). Helium ion microscopy for high-resolution visualization of the articular cartilage collagen network. J Microsc 246, 168–176.
Wagner CA, Finberg KE, Breton S, Marshansky V, Brown D & Geibel JP (2004). Renal vacuolar H+-ATPase. Physiol Rev 84, 1263–1314.
Ward B, Notte JA & Economou NP (2007). Helium-ion microscopy. Photonics Spectra 41, 68–70.