
Physiology News Magazine

Muscle afferents, back again?
James Fisher and Michael White explain how afferent feedback from the exercising muscle can be modified in health and disease – and how recent research is helping us to understand the neurophysiological basis for these in vivo observations
Features
Muscle afferents, back again?
James Fisher and Michael White explain how afferent feedback from the exercising muscle can be modified in health and disease – and how recent research is helping us to understand the neurophysiological basis for these in vivo observations
Features
James P Fisher & Michael J White
School of Sport and Exercise Sciences, University of Birmingham
https://doi.org/10.36866/pn.59.35


Classically, the cardiovascular response to exercise is said to be governed by both central feed-forward and peripheral feedback mechanisms. During exercise, slowly conducting group III and IV muscle afferents are activated by mechanical and metabolic stresses placed on the muscle. These muscle afferents relay feedback to the cardiovascular areas of the brain stem and cause both an inhibition of vagal outflow and an excitation of the sympathetic nervous system. This feedback, together with central command, produces the increase in heart rate and blood pressure associated with exercise (Fig. 1).

Activation of muscle afferents is intimately involved in resetting and adjusting the sensitivity of the baroreflex during exercise. Whilst muscle metaboreceptor activation is known to increase baroreflex sensitivity, muscle mechanoreceptor activation decreases the sensitivity of the baroreflex (Carrington & White, 2001). An often overlooked effect of muscle afferent activation is the modulation of motor neurone excitability in the spinal cord, a factor which has major implications for those interested in the mechanisms of central fatigue (see Gandevia, 2001 for review).
Importantly, afferent feedback from the active muscle can be modified by factors other than simply exercise intensity, for example muscle fibre type, training status, mass and temperature (Fig. 2).
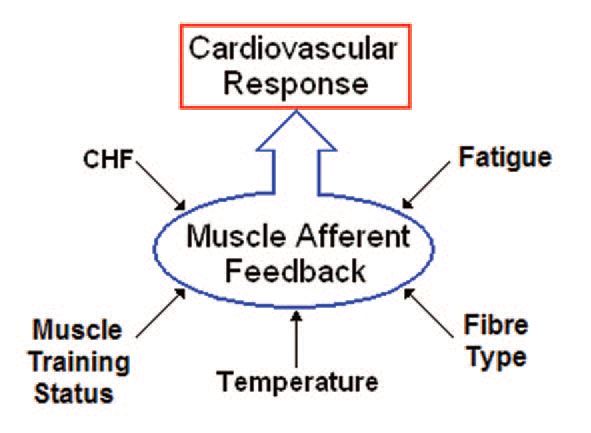
Studies in both animals and humans have shown that the pressor response, resulting from isometric exercise of muscle with a faster contractile character and isomyosin content, is greater than that from a muscle of slower contractile character. This is because, for a given level of force production, fast fibres produce a greater metabolic stimulus to the muscle afferents. However, athletes who undertake training programs that place a high anaerobic load on specific muscle groups appear to attenuate this stimulus and have a smaller than usual pressor response to electrically evoked exercise of those trained muscle groups (Carrington et al. 1999). During electrically-evoked exercise there is no central command, so reduction of the exercise pressor reflex clearly indicates an attenuated afferent feedback. Similarly, longitudinal laboratory-based studies have shown that specific local muscle training also blunts the cardiovascular response to isometric exercise (Fisher& White, 1999). Thus it appears that training attenuates the stimulation of muscle afferents, either by reducing metabolite accumulation, or in some instances by blunting their sensitivity through chronic exposure to the products of anaerobic metabolism.
There may be surprising parallels between the local muscle conditions experienced by athletes training for longer sprint events (i.e. 400 m) and by the low-flow conditions in, for example, the muscles of chronic heart failure (CHF) patients. This might explain some similarities in muscle afferent-driven cardiovascular responses to exercise of their muscles. However even within the same muscle responses to mechanical or metabolic stress may be changed differentially due to training or disease. For example, Sterns et al. (1991) demonstrated that, when the muscle metaboreflex is activated in isolation by occluding the circulation to the forearm immediately after exercise, CHF patients show attenuated muscle sympathetic nerve activity compared with matched controls. As pH was found to be similar at this time, this suggests a desensitisation of the muscle metaboreflex. Furthermore, as muscle sympathetic nerve activity increased normally during exercise in CHF patients, there must have been an exaggerated contribution from either the muscle mechanoreflex or central command.
This complex situation may have been clarified in a recent study by Li et al. (2004) which suggests differential change in the sensitivity of specific receptors on muscle metaboreceptors (VR1) and muscle mechanoreceptors (P2X) in an animal model of heart failure. Whilst the sensitivity of VR1 receptors was blunted in heart failure, the sensitivity of P2X receptors was augmented. This is a major step forward in our understanding, as the properties of these receptors are well documented in vitro. For example, the activity of the VR1 receptor normally increases as pH falls and temperature rises. Of course, both of these changes occur in muscle during isometric or dynamic activity if the intensity is high enough or if blood flow is restricted.
How these receptors are affected in human muscle which has become adapted, or is undergoing adaptation, to altered activity pattern or disease is evidently an important question and one requiring an integrative physiological approach. It seems that – forgive the pun – the story of muscle afferents in human exercise physiology still has a long way yet to run.
Acknowledgements
JPF is kindly funded by the British Heart Foundation.
References
Carrington CA & White MJ (2001). Exercise induced muscle chemoreflex modulation of spontaneous baroreflex sensitivity in man. J Physiol 536, 957-62.
Carrington CA, Fisher WJ, & White MJ (1999). The effects of athletic training and muscle contractile character on the pressor response to isometric exercise of the human triceps surae. Eur J Appl Physiol 80, 337-343.
Fisher WJ & White MJ (1999). Training-induced adaptations in the central command and peripheral reflex components of the pressor response to isometric exercise of the human triceps surae. J Physiol 520, 621-628.
Fisher JP & White MJ (2004). Muscle afferent contributions to the cardiovascular response to isometric exercise. Exp Physiol 89, 639646.
Gandevia S.C. (2001). Spinal and supraspinal factors in human muscle fatigue. Physiol Rev 81, 1725-1789.
Li J., Sinoway A.N., Gao Z., Maile M.D., Pu M. and Sinoway L.I (2004). Muscle mechanoreflex and metaboreflex responses after myocardial infarction in rats. Circ 110, 3049-3054.
Sterns DA, Ettinger SM, Gray KS, Whisler SK, Mosher TJ, Smith M, & Sinoway L (1991). Skeletal muscle metaboreceptor exercise responses are attenuated in heart failure. Circ 84, 2034-2039.