
Physiology News Magazine

My top ten papers
Jonathan Ashmore reflects on the papers that have most influenced his career
Membership
My top ten papers
Jonathan Ashmore reflects on the papers that have most influenced his career
Membership
https://doi.org/10.36866/pn.88.44

I started my scientific career as a theoretical physicist. That phase didn’t last long. Real hands-on lab work was easily much more fun. This history explains the first 2.5 papers. There is naturally a biophysical slant to the others, although I would have liked to have included many more. Disloyally, some of the later papers are not taken from J Physiol, a trend in neuroscience which I hope The Journal can reverse.
1
Dyson FJ (1949). The S matrix in quantum electrodynamics. Phys Rev 75, 1736–1755.
As theoretical physics graduate students, we actually read alarmingly few papers: the information came through precirculated
‘preprints’ appearing often years before the journal version. But this paper we read for inspiration: it outlines a programme of how to remove annoying infinities in the calculations (the denominator was often 0!) from an otherwise beautiful theory. The problem of renormalization is still with us in the post-Higgs world, but this was my introduction to a complex edifice where, as in physiology, the devil is in the detail.
2
Marr DA (1969). Theory of cerebellar cortex. J Physiol 202, 437–470.
This paper, written by David Marr when he was still a PhD student with Giles Brindley, combines an elegant theoretical idea about how to extract complex patterns from an even more elegant structure, the cerebellum, and form a wide range of complex motor responses. As a theory paper it had a remarkable impact just at the moment when LTP was just about to be discovered.
3
Hodgkin AL & Huxley AF (1952).
A quantitative description of membrane current and its application to conduction and excitation in nerve. J Physiol 117, 500–544.
Published exactly 60 years ago, this paper is still just as fresh today as it was then. But, encountered with the mindset of another discipline the description is hard to stomach – where do the ideas come from? It is really a ‘phenomenological’ model (i.e. an informed way of fitting data), but with a long tradition of physiology. The effort to compute the correct action potential velocity, all done by Andrew Huxley using just a hand-cranked Brunsviga calculator, today makes the blood run cold.
4
Fatt P & Katz B (1952). Spontaneous subthreshold activity at motor nerve endings. J Physiol 117, 109–128.
1952 must count as an annus mirabilis of The Journal. This paper describes the discovery (‘a chance observation’) of miniature endplate potentials in the early intracellular recordings with microelectrodes. It expresses all the excitement of finding something completely new and the possible explanations are discussed with an incisive logic. With hindsight, many of the observations, (for example, that the frequency but not the amplitude of the minis is calcium-dependent), is the foundation of much subsequent synaptic physiology.
5
Hamill OP, Marty A, Neher E, Sakmann B & Sigworth FJ (1981). Improved patch-clamp techniques for high-resolution current recording from cells and cell-free membrane patches. Pflugers Arch 391, 85-100.
This is the methods paper par-excellence. It let patch-clamping become one of the key enabling technologies of neuroscience. It has often (maliciously) been said that, had patch-clamping been discovered in a molecular biology lab, we would still not know how to do it!
6
Corey DP & Hudspeth AJ (1979). Response latency of vertebrate hair cells. Biophys J 26, 499–506.
This paper firmly established the notion that hair cell transduction is ‘direct’. A chemical signal between bending the stereocilia and the opening of the ionic channel was a hypothesis which until then was at least plausible given what was then known about photoreceptors. It opened up the problem of identifying the molecular basis of sound transduction. The question is still un-answered.
7
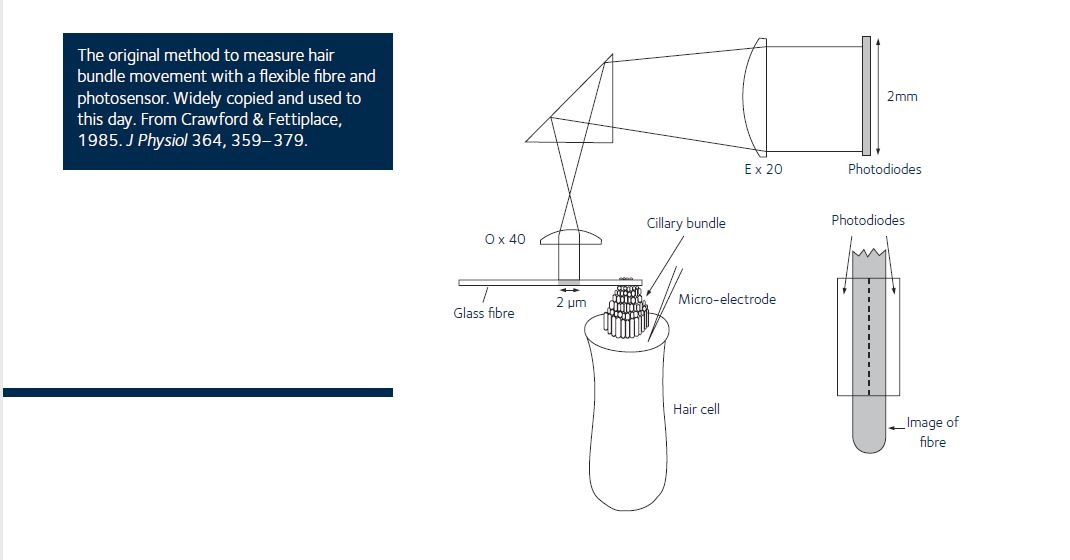
Crawford AC & Fettiplace R (1985). The mechanical properties of ciliary bundles of turtle cochlear hair cells. J Physiol 364, 359–379.
The first convincing observations of ‘active’ processes in hearing. The experiments describe nanometre movements in hair bundles, but in non-mammalian hearing organs. These results spawned a whole research field which is still thriving today and fuelled the debate about what amplifies sound in the mammal (which is categorically not, in my view, the hair bundle!).
8
Hilgemann DW (1994). Channel-like function of the Na+,K+ pump probed at microsecond resolution in giant membrane patches. Science 263, 1429–1432.
This is another technically inspirational paper. Don Hilgemann rebuilt an Axon amplifier to record ultrafast events in membrane biophysics, the transition of ions into the sodium pump. Not only that, it addressed the perennial problem of how to isolate very small amounts of the pump in membranes. I like this paper because it has a definite ‘wow!’ factor.
9
Zheng J, Shen W, He DZ, Long KB, Madison LD & Dallos P (2000). Prestin is the motor protein of cochlear outer hair cells. Nature 405, 149–155.
This paper identified the key motor molecule that makes outer hair cells move. Jing Zheng, as a side-project in Peter Dallos’s lab, had been collecting enough inner and outer hair cells over 12 months (by aspirating single cells), to make a cDNA subtraction library. Although the paper does contain a suspect calibration, the effect on the field of identifying the molecule was profound. We have yet to work out prestin’s structure.
10
Goutman JD & Glowatzki E (2007). Time course and calcium dependence of transmitter release at a single ribbon synapse. Proc Natl Acad Sci U S A 104, 16341–16346.
This paper describes experiments where both presynaptic and the (extremely small) postsynaptic bouton of the hair cell afferent dendrite were simultaneously patch-clamped. It demonstrates convincingly the hair trigger release of glutamatergic vesicles, probably as multi-vesicular events, at the ribbon synapse. Not only is it a technical tour-de-force (it describes those experiments that only work once every month) but it describes the results with impeccable economy.
It might be said that I have chosen too many ‘old’ papers. To which I reply that these papers were indeed formative – they have a fine balance of numeracy and description – and, even now, on re-reading them, I think that they have stood the test of time well as models of how to present good and exciting science.