
Physiology News Magazine

Sympathetic nerve activity and cardiac output: an integrated balance with implications for blood pressure regulation
Sympathetic vasoconstrictor nerves have an important role in blood pressure regulation in humans via control of peripheral vascular resistance. Recent findings of a balance between cardiac hemodynamics and control of sympathetic activity at rest provide insight into integrated mechanisms of blood pressure regulation in individuals with widely varying sympathetic activity
Features
Sympathetic nerve activity and cardiac output: an integrated balance with implications for blood pressure regulation
Sympathetic vasoconstrictor nerves have an important role in blood pressure regulation in humans via control of peripheral vascular resistance. Recent findings of a balance between cardiac hemodynamics and control of sympathetic activity at rest provide insight into integrated mechanisms of blood pressure regulation in individuals with widely varying sympathetic activity
Features
Nisha Charkoudian, Michael J Joyner, & B Gunnar Wallin
Departments of Physiology and Anesthesiology, Mayo Clinic College of Medicine, Rochester, MN, USA and Institute of Clinical Neuroscience, Göteborg University, Göteborg, Sweden
https://doi.org/10.36866/pn.62.25

Sympathetic vasoconstrictor neural activity (SNA) is a major contributor to blood pressure regulation in humans: a rise in SNA increases vasoconstriction and vascular resistance, resulting in an increase in blood pressure. It has long been considered paradoxical, therefore, that humans with higher resting SNA do not necessarily have higher blood pressures than those in whom SNA is very low. Indeed, there is wide interindividual variability in SNA among normotensive humans with similar arterial pressures. Furthermore, muscle SNA (MSNA, often measured in humans) is extremely reproducible over time (Fagius & Wallin, 1993), and correlates well with resting SNA in other vascular beds (Wallin et al. 1996). Yet MSNA can vary 5-10 fold among individuals (Fagius & Wallin, 1993; Charkoudian et al. 2005). Thus if we consider that mean arterial pressure is determined by the product of cardiac output and total peripheral resistance (MAP = CO x TPR), we must ask the question: how can all this variability exist in normal healthy individuals with similar blood pressures?
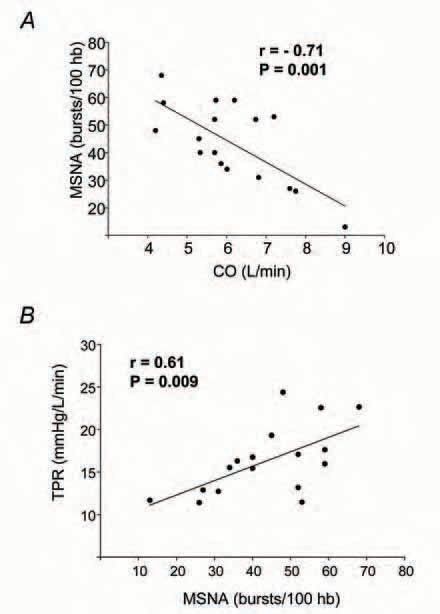
We recently set out to investigate this apparent paradox by testing the hypothesis that a balance exists between CO and MSNA that is important to normal blood pressure regulation. We further hypothesized that this balance would involve an inhibitory influence of CO on baroreflex control of MSNA. To address these hypotheses, we measured MSNA (peroneal microneurography), CO (acetylene uptake) and arterial pressure (directly via radial artery catheter) in 18 normotensive men (Charkoudian et al. 2005). Our most striking finding was a strong inverse relationship between CO and MSNA, such that individuals with high CO tended to have low mean levels of MSNA, and vice versa. Figure 1A shows the relationship between CO and MSNA at rest among our subjects. Furthermore, MSNA showed a positive correlation with TPR in our subjects, as shown in Figure 1B. Taken together, these relationships suggested a balance between the two main contributors to arterial pressure: CO and TPR (Charkoudian et al. 2005). The findings provide strong evidence that a major reason why individuals with higher sympathetic nerve activity do not have higher blood pressures is because of a balance between cardiac hemodynamics and control of peripheral vascular resistance.
Previous microneurographic recordings of human MSNA have revealed that, in individual subjects, the arterial baroreflex exerts substantial control over MSNA. Thus small changes in blood pressure elicit marked opposing changes in MSNA, which tend to correct, or ‘buffer’, the original changes in pressure (Kienbaum et al. 2001; Charkoudian et al. 2004). Given this strong baroreflex control, it is quite striking that the large inter-individual variability in MSNA in normotensive humans results in a complete lack of correlation between mean levels of nerve activity and blood pressure across individuals (Skarphedinsson et al. 1997; Charkoudian et al. 2005). In our recent study, we reasoned that this lack of correlation might be explained by an influence of CO on baroreflex control of MSNA (Charkoudian et al. 2005). We assessed this possibility using a technique called ‘threshold analysis’, which gives information about how likely it is that a given level of blood pressure will elicit a burst of sympathetic activity. The midpoint (T50) of a threshold diagram provides a measure of the blood pressure value which elicits a burst of sympathetic activity 50% of the time (for detailed discussion, see Kienbaum et al. 2001). We found that CO and stroke volume (SV) were each inversely related to T50 among our subjects, suggesting that high resting CO and SV may inhibit sympathetic outflow at rest via the baroreflex.
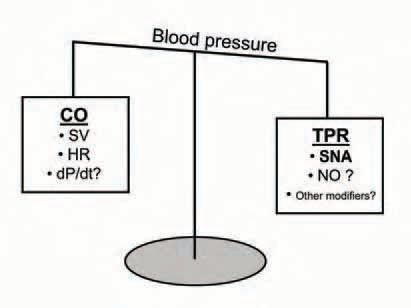
The inverse relationship we observed between CO and MSNA, and the apparent inhibitory influences of CO and SV on baroreflex control of MSNA, suggest that a balance between CO and SNA is an important element of normal blood pressure regulation. This is shown schematically in Fig. 2, along with factors that may influence this balance. From the cardiac perspective, the influence of SV may be important, via afferents from the cardiopulmonary region (Charkoudian et al. 2004), and/or by its effects on the rate of change of pressure (dP/dt) which could alter afferent firing of arterial baroreceptors (Chapleau & Abboud, 1989). The relationship between SNA and total peripheral resistance (TPR) in this balance may be affected by one or more of numerous influences on vascular tone. For example, Skarphedinsson et al. (1997) reported a direct relationship between circulating nitrite/nitrate (a plasma marker of nitric oxide activity) and resting MSNA among normotensive males. This suggests that individuals with higher SNA also have higher circulating nitric oxide, and that its vasodilator effects may offset vasoconstrictor influences of high sympathetic activity.
Our recent results do not address the origin of the inter-individual variability in MSNA or CO. Some evidence suggests that the variability in the MSNA/CO relationship may be genetic in origin. For example, identical twins have very similar levels of resting MSNA (Wallin et al. 1993) and genetic variation in genes encoding the betaadrenergic receptor cause differences in CO and hemodynamic responses (Eisenach et al. 2005). It will be interesting to see whether it is possible in the future to identify genetic factors which contribute specifically to the balance of factors described here.
The existence of a balance between CO and MSNA leads to the intriguing possibility that the bases for some forms of hypertension may be that individuals are ‘out of balance’ in this regard – their level of CO is too high for the level of MSNA, or vice-versa. Taking both elements of the CO-MSNA balance into account may help to explain previously contradictory reports regarding whether individuals with hypertension have higher SNA compared to healthy controls (Gudbjornsdottir et al. 1996). In a broader sense, better understanding of the integrated balance of the mechanisms presented here (and other potential contributors) will not only improve our understanding of the complex physiology involved, but may, in the future, lead to more individualized diagnostic assessment and treatment of certain forms of cardiovascular malfunction.
References
Chapleau MW & Abboud FM (1989). Determinants of sensitization of carotid baroreceptors by pulsatile pressure in dogs. Circ Res 65, 566-577.
Charkoudian N, Joyner MJ, Johnson CP, Eisenach JH, Dietz NM & Wallin BG (2005). Balance between cardiac output and sympathetic nerve activity in resting humans: role in arterial pressure regulation. J Physiol 568, 315-321.
Charkoudian N, Martin EA, Dinenno FA, Eisenach JH, Dietz NM & Joyner MJ (2004). Influence of increased central venous pressure on baroreflex control of sympathetic activity in humans. Am J Physiol Heart Circ Physiol 287, H1658-1662.
Eisenach JH, Barnes SA, Pike TL, Sokolnicki LA, Masuki S, Dietz NM, Rehfeldt KH, Turner ST & Joyner MJ (2005). Arg16/Gly {beta}2adrenergic receptor polymorphism alters the cardiac output response to isometric exercise. J Appl Physiol 99, 1776-1781.
Fagius J & Wallin BG (1993). Long-term variability and reproducibility of resting human muscle nerve sympathetic activity at rest, as reassessed after a decade. Clin Auton Res 3, 201-205.
Gudbjornsdottir S, Lonnroth P, Sverrisdottir YB, Wallin BG & Elam M (1996). Sympathetic nerve activity and insulin in obese normotensive and hypertensive men. Hypertension 27, 276-280.
Kienbaum P, Karlssonn T, Sverrisdottir YB, Elam M & Wallin BG (2001). Two sites for modulation of human sympathetic activity by arterial baroreceptors? J Physiol 531, 861-869.
Skarphedinsson JO, Elam M, Jungersten L & Wallin BG (1997). Sympathetic nerve traffic correlates with the release of nitric oxide in humans: implications for blood pressure control. J Physiol 501, 671-675.
Wallin BG, Kunimoto MM & Sellgren J (1993). Possible genetic influence on the strength of human muscle nerve sympathetic activity at rest. Hypertension 22, 282-284.
Wallin BG, Thompson JM, Jennings GL & Esler MD (1996). Renal noradrenaline spillover correlates with muscle sympathetic activity in humans. J Physiol 491, 881-887.