
Physiology News Magazine
The physiology of Casino Royale: proteins to parkour
Features
The physiology of Casino Royale: proteins to parkour
Features
Martin McDonagh
School of Sport and Exercise Sciences, University of Birmingham, UK
https://doi.org/10.36866/pn.81.29

In the film Casino Royale, James Bond chases terrorist Mollaka over a construction site in Madagascar. Mollaka leaps from crane to rooftop and from rooftop to ground, landing under control and able to resume running immediately. These stunts are not just filmic tricks – Mollaka (Sébastien Foucan) is a top class exponent of free running or parkour (see www.youtube.com/watch?v=Q0e7akuSgKI ). Despite Foucan’s skill the shock to the legs is still enormous – 17 body weights for a 1.8 metre fall (Yeadon et al. 2010). How can he absorb this shock without injury? In engineering, shock absorbers prevent high-impact forces by creating constant deceleration over a given distance or stroke length. Springs produce high decelerations towards the end of the impact and dashpots produce high decelerations at the start, thus many industrial shock absorbers use a combination of the two.
So where are the springs and dashpots of human shock absorbers and how do they work? To answer this we need to integrate research findings all the way from the molecular mechanisms of muscle cells to the neural control of falling and landing.
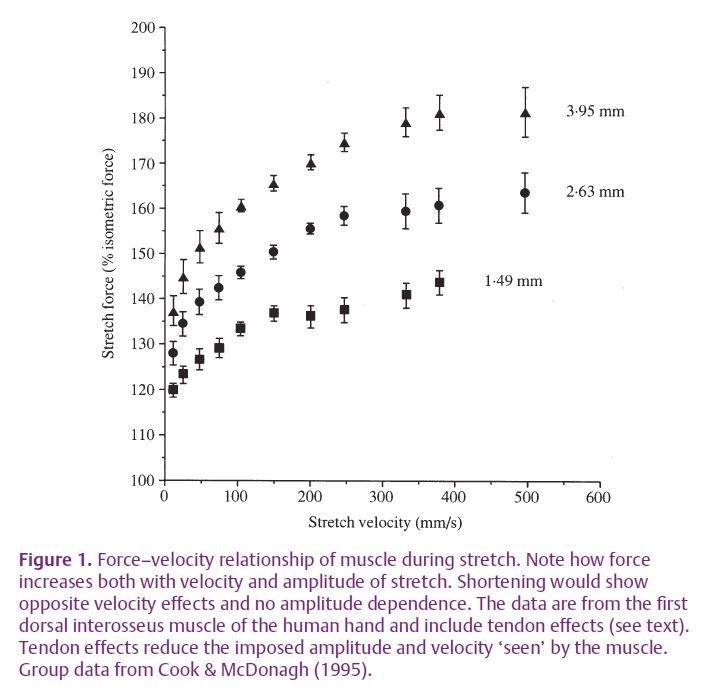
On landing the impact causes ankle, knee and hip joints to rotate. This stretches the muscles opposing the movement which lengthen under control, reducing joint rotations and acting as a brake. When muscles are braking and lengthening the relationship between force and velocity is quite different from A.V. Hill’s classic shortening force–velocity curve. Firstly, force increases with velocity (dashpot effect); secondly, force increases with the amplitude of stretch (spring effect); and thirdly, the force can be up to twice that produced by an isometric contraction. Figure 1 shows the combined effects of amplitude and velocity for a human muscle. These properties make stretching muscle a particularly effective brake; the greater the shock the greater the force response. So what are the mechanisms of these properties at the molecular level?
Muscle force increases with the velocity because stretch causes high positive strain in muscle cross-bridges. This slows their power stroke leaving more and more pre-stroke bridges attached, as velocity increases (Pinniger et al. 2006a). Force increases with stretch amplitude because stretch increases the strain in the elastic crossbridges considerably above the values found in an isometric contraction. The force rises in proportion to the strain until bridge detachment limits the maximum force to around twice the isometric value. Furthermore, recent work has also suggested that greater stretch favours the attachment of the second myosin head found on all muscle crossbridges ( Brunello et al. 2007) thus increasing the available force. Elastic proteins such as titin within the sarcomere and transverse elastic connections between myofibrils may also increase force when stretched (Pinniger et al. 2006b).
Muscle fibres, although beautifully evolved to act as brakes, do not act alone. The shock of landing is also absorbed by the tendons to which the muscles are attached. Tendons have a complex structure but are mainly composed of the elastic protein collagen. They have 70 times the Young’s Modulus of fully activated muscle, but their stiffness relative to muscle is reduced because their cross-sectional area is often 30–100 times smaller than the attached muscle. With lower stiffness the tendon can provide a greater proportion of the total lengthening of the muscle–tendon complex. Millimetres of the tendon lengthening will be particularly large when the tendon is long and thin and the muscle short and fat. For example, it can be calculated from known values of Achilles tendon stiffness and peak impact forces that this tendon could stretch by 13 mm or 16% following a fall from 0.8 m. Tendon stretch has the advantage of both increasing the absorption of energy and preventing over-lengthening and damage to muscle fibres (Cook & McDonagh, 1995).
The energy of impact must be dissipated as well as absorbed, otherwise on landing the subject will be thrown upwards again by elastic recoil. Energy is dissipated as heat by the muscle component which acts as a damper as well as a spring. Damping occurs when impact forces cause actin and myosin filaments to slide apart, detaching crossbridges as they go. In summary, the muscle–tendon complexes of the body are a multi-mass, damped-spring suspension system.
The absorption and dissipation of energy is sufficient to brake a fall because all the joints of the leg are involved. The sum of their rotation amplitudes (e.g. 33 deg ankle, 66 deg knee, 54 deg hip for a 1 m fall) determines the distance the centre of gravity of the body moves following impact. This is equivalent to the stroke length of a mechanical shock absorber. In simple terms F = m g h/∆h, with gravitational acceleration (g), the longer the stroke length (∆h), the smaller the average force (F) experienced over the stroke length by the centre of gravity of the subject’s mass (m) and the smoother the landing from height (h) ( Minetti et al. 1998). Joint rotations and centre of gravity excursions increase with higher falls (Santello et al. 2001). In parkour the impact force is further reduced by a roll on landing which increases the distance and surface area over which the shock is absorbed. Compression shock to bones on impact is also minimized by landing with slightly flexed joints which rapidly rotate. In addition, co-contraction of the flexors ensures correct stiffness of the extensor muscle brakes combined with the appropriate joint angle prior to impact (Yeadon, 2010)
Muscle–tendon complexes are more than just passive suspension systems, as their damping and the spring constants can be modulated by varying the activation of the muscle component. This is an active suspension system, exactly the type of suspension banned by Formula 1! With too little activation the joints over rotate and the subject collapses in a heap and cannot continue running. With too much activation muscles are stiff and the subject lands like a steel pin, joints under rotate, impact forces are high and bones may be broken. How does the brain get the activation right?
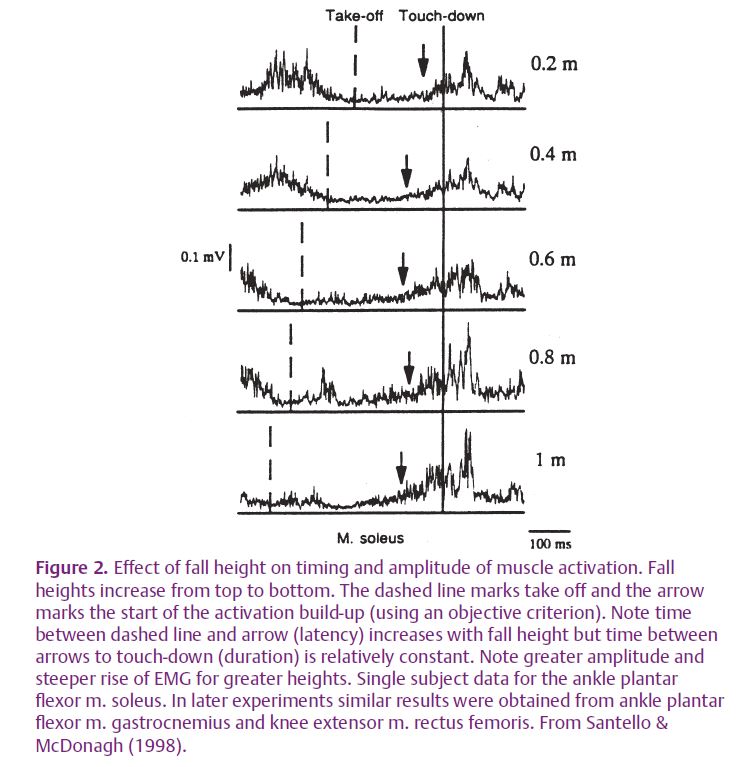
To tackle this question we varied the impact force by asking subjects to fall from different heights whilst we recorded muscle activations from several flexor and extensor muscles of the leg (Santello et al. 2001). Figure 2 shows the results. The further the subjects fell the greater the activation of the muscles during the flight. Thus, the central nervous system (CNS) predicts the required activation for each height rather than setting one activation level for all heights. The brain also needs to predict the timing of the impact. A simple strategy would be to contract all the muscles at the same delay after take-off. Is this what happens? No, as Fig. 2 shows, the longer the fall, the longer the brain waits before triggering the contraction. However, the duration of the next time period, from the start of muscle activation to floor contact, is relatively constant, irrespective of fall height. This period also shows how the motor programme increases force. As greater fall heights are perceived, activation rises more steeply so matching muscle force to the expected impact. In summary, the brain appears to predict both the force and timing of impact and to pre-programme the muscle activation required to deal with it. The mechanisms behind these predictions require much further investigation but probably involve a combination of visual distance estimation, mental clocks and sensorimotor memories.
However, prediction is not the whole story. An examination of Fig. 2 shows that EMG activity continues beyond impact. Is this a continuation of the pre-programmed activation or a reflex triggered by the impact? To settle this question we tricked the brain into expecting a landing after a short fall (0.45 m) but actually provided a lower landing surface at 0.7 m (Duncan & McDonagh, 2000). Subjects expected to land on a false floor which they saw 0.45 m below the take-off platform. This floor easily gave way on contact with joint rotation and muscle stretch only occurring when the subject hit the hard floor a further 0.25 m below. Falls to visible hard floors at 0.45 m and 0.7 m provided controls. The results were clear. The reflex was only present if joints rotated. Figure 3 shows how the false floor trials separated the pre-programmed from the reflex components. This is particularly obvious for the gastrocnemius muscle. Muscle activation whilst airborne was clearly timed to the impact with the false floor. This activation then died away as the subject descended below the false floor to impact on the hard floor. On impact, joints rotated and a reflex response occurred at a mean latency of 56 ms. Thus, the control of landing depends on both pre-programmed and stretch reflex responses.
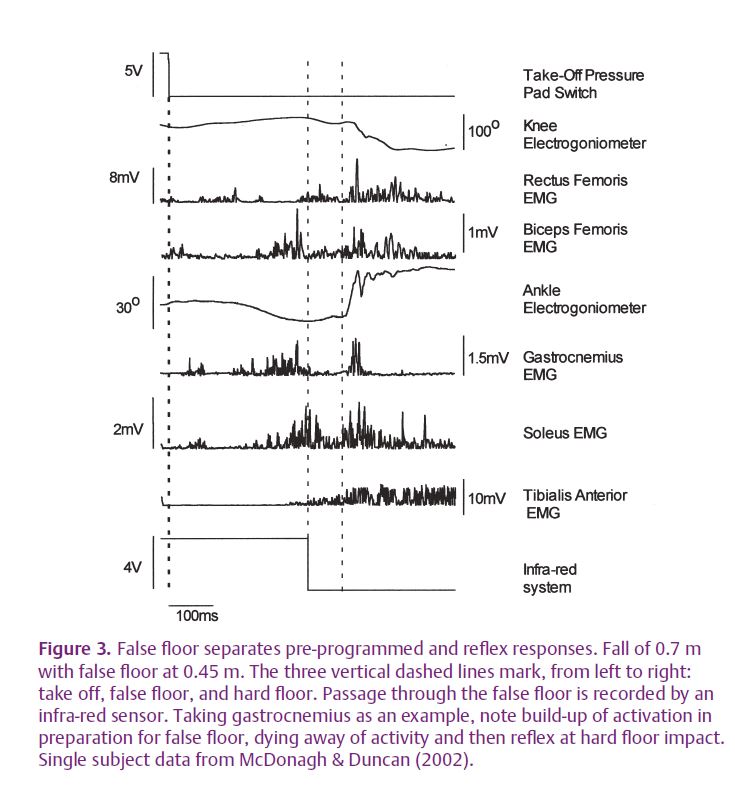
Our data had one further surprise in store. The amplitude of the reflex at hard floor impact in the ‘via false floor’ trials was double that for the direct fall (McDonagh & Duncan, 2002). However, joint rotations at impact were the same in each condition and there was no joint rotation at false floor level. Thus, there was a larger response for the same stimulus i.e. an increase in gain of the reflex loop. Eventually we concluded that this increase in gain was triggered by the non-occurrence of expected joint rotation at the predicted time of impact i.e. at the false floor. In other words, the absence of an expected event is as much a signal for the CNS as the presence of an unexpected one. This idea is probably widely true in many other CNS control systems and deserves much further investigation.
We can now answer, in a nutshell, the ‘Casino Royale’ question. Sébastien Foucan can absorb the shock of parkour because muscle–tendon complexes have evolved many ingenious shock-absorbing proteins, and mechanisms. The brain then tunes these shock absorbers using both pre-programmed control and adjustable reflexes to achieve injury-free landings over a wide range of impact conditions.
References
Brunello E, Reconditi M, Elangovan R, Linari M, Sun Y, Narayanan T, Panine P, Piazzessi G, Irving M & Lombardi V (2007). Skeletal muscle resists stretch by rapid biding of the second motor domain of myosin to actin. Proc Natl Acad Sci U S A 114, 20115–20119.
Cook CS & McDonagh MJN (1995). Force responses to controlled stretches of electrically stimulated human muscle–tendon complex. Exp Physiol 80, 477–490.
Duncan A & McDonagh MJN (2000). Stretch reflex distinguished from pre-programmed muscle activations following landing impacts in man. J Physiol 526, 457–468.
McDonagh MJN & Duncan A (2002). Interaction of pre-programmed control and natural stretch reflexes in human landing movements. J Physiol 544, 985–994.
Minetti AE, Ardigo LP, Susta D & Cotelli F (1998). Using leg muscles as shock absorbers; theoretical predictions and experimental results of drop landing. performance. Ergonomics 41, 1771–1791.
Pinniger G, Ranatunga KW & Offer G (2006a). What structures bear the tension and store energy in lengthening muscle? Physiology News 65, 1–2.
Pinniger G, Ranatunga KW & Offer G (2006b). Crossbridge and non-crossbridge contributions to tension in lengthening rat muscle: force-induced reversal of power stroke. J Physiol 573, 627–643.
Santello M & McDonagh MJN (1998). The control of timing and amplitude of EMG activity in landing movements in humans. Exp Physiol 83, 857–874.
Santello M, McDonagh MJN & Challis JH (2001). Visual and non-visual control of landing movements in humans. J Physiol 537, 313–327.
Yeadon NR, King MA, Forrester SE, Caldwell GE & Pain MTG (2010). The need for muscle co-contraction prior to landing. J Biomech 43, 364–369.