
Physiology News Magazine
Two arms as one: position and movement sense in an elbow-matching task
Masahiko Izumizaki and Uwe Proske have found that when subjects match the position of one unseen arm by placement of the other, sensory information coming from both arms is used to achieve the observed matching accuracy.
Features
Two arms as one: position and movement sense in an elbow-matching task
Masahiko Izumizaki and Uwe Proske have found that when subjects match the position of one unseen arm by placement of the other, sensory information coming from both arms is used to achieve the observed matching accuracy.
Features
Masahiko Izumizaki (1) and Uwe Proske (2)
1: Department of Physiology, Showa University School of Medicine, Tokyo, Japan
2: Department of Physiology, Monash University, Clayton, Victoria, Australia
https://doi.org/10.36866/pn.82.26


This story is about the kinaesthetic senses. They are part of a group of senses, the proprioceptive senses, concerned with signalling the body’s own actions. The kinaesthetic senses comprise the sense of limb position and the sense of limb movement (Proske & Gandevia, 2009). Other proprioceptive senses include the senses of effort, tension and balance. The proprioceptive senses are somewhat mysterious because we are largely unaware of them. When we look at an object or touch it we experience a specific, identifiable sensation. By contrast, we know precisely where our limbs are when we are not looking at them, but that awareness is not accompanied by any identifiable sensation. It presumably means that the sensory information concerned with signalling the position and movement of our limbs is processed largely unconsciously.
The present day view is that the principal kinaesthetic sensors are the muscle spindles. At some joints, particularly the finger joints, skin and joint receptors also contribute to the senses of position and movement. At the elbow joint skin stretch receptors are likely to supplement the main kinaesthetic signal coming from spindles in elbow muscles. The primary endings of muscle spindles are thought to contribute to both the sense of limb position and the sense of limb movement while secondary endings of spindles contribute to the sense of position (McCloskey, 1973).
We have been studying position sense at the forearm in a simple matching task. One arm, designated the reference arm, is placed in a particular position and the blindfolded subject is asked to match it with their other, indicator arm. Subjects are normally quite accurate and are able to match positions with errors of 2 deg or less. If, instead of using a matching process, subjects are asked to indicate the position of one arm with a pointer, they are less accurate. It has led us to conclude that arm matching is a cooperative process in which both arms play a contributory role. In order to further explore this idea we have done some simple experimental manipulations of one arm to see what effect it had on matching accuracy (Izumizaki et al. 2010). We carried out these matches under two conditions, one where the reference arm lay relaxed on a support and the second, when its elbow flexor muscles were vibrated at 80 Hz. Muscle vibration is a largely selective stimulus for the primary endings of muscle spindles. The extra impulses generated by the vibration lead to an illusion of limb movement and displaced position (Goodwin et al. 1972). We wanted to know whether altering the conditions of the indicator arm affected matching accuracy when the reference arm lay relaxed and when it was being vibrated, that is, in the presence of a large, false proprioceptive signal coming from the reference arm.
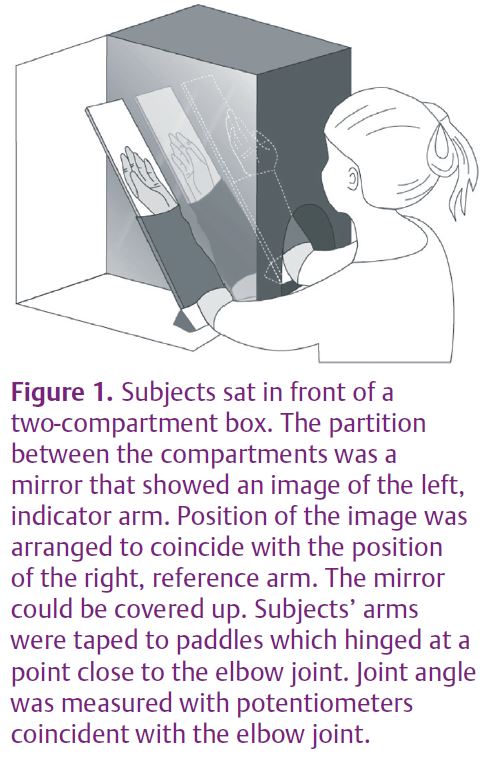
The manipulations of the indicator arm were of four different kinds. In all trials the reference arm lay hidden from the subject’s view. In the first series of matches the experiment was done in the traditional way, with the subject blindfolded so that they could see neither arm. In the second series subjects could see their indicator arm; in the third the indicator was placed out of view and subjects were required to match the perceived position of the reference arm by placement of a dummy arm. Finally, a mirror was slipped into the partition between the two arms (Fig. 1). The position of the mirror was arranged so that the reflected image of the indicator arm appeared to occupy a position coincident with that of the reference arm. Here subjects were instructed to look only at the mirror image and not at the actual indicator. Example records of matching trials are shown in Fig. 2 and the group results are shown in Fig. 3.
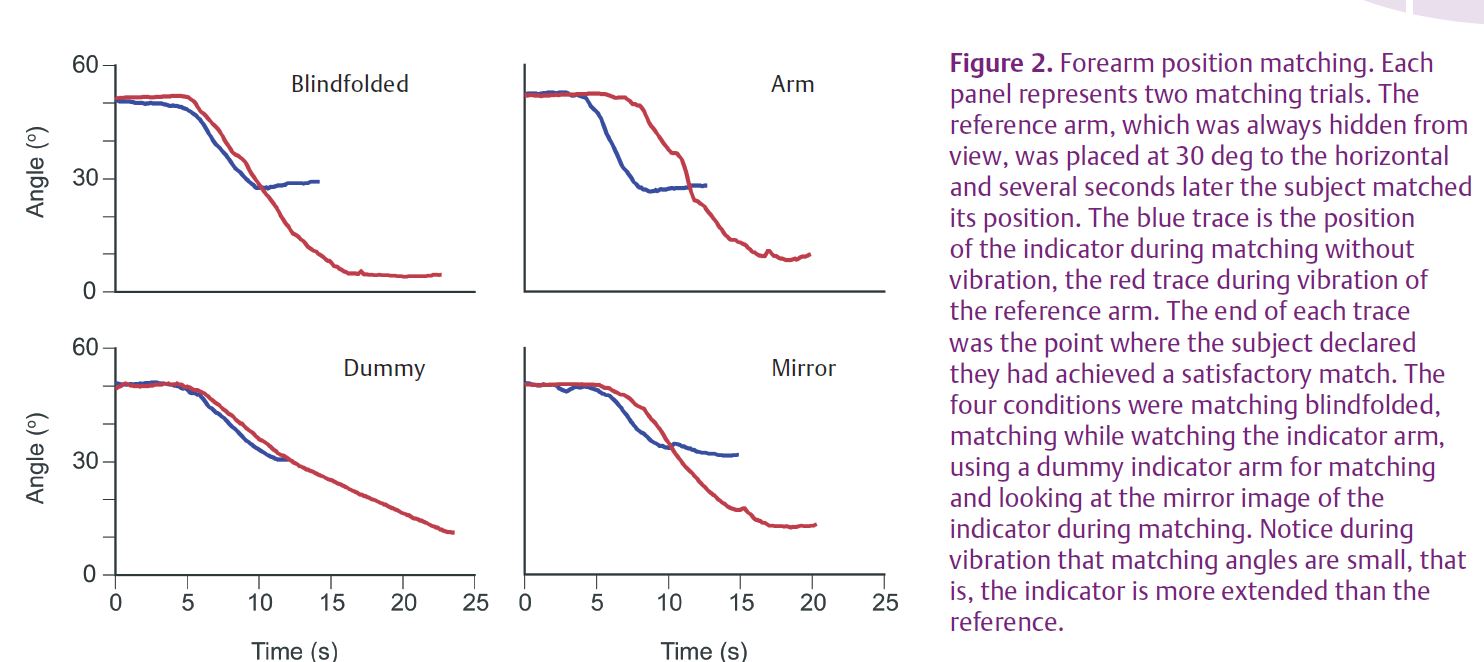
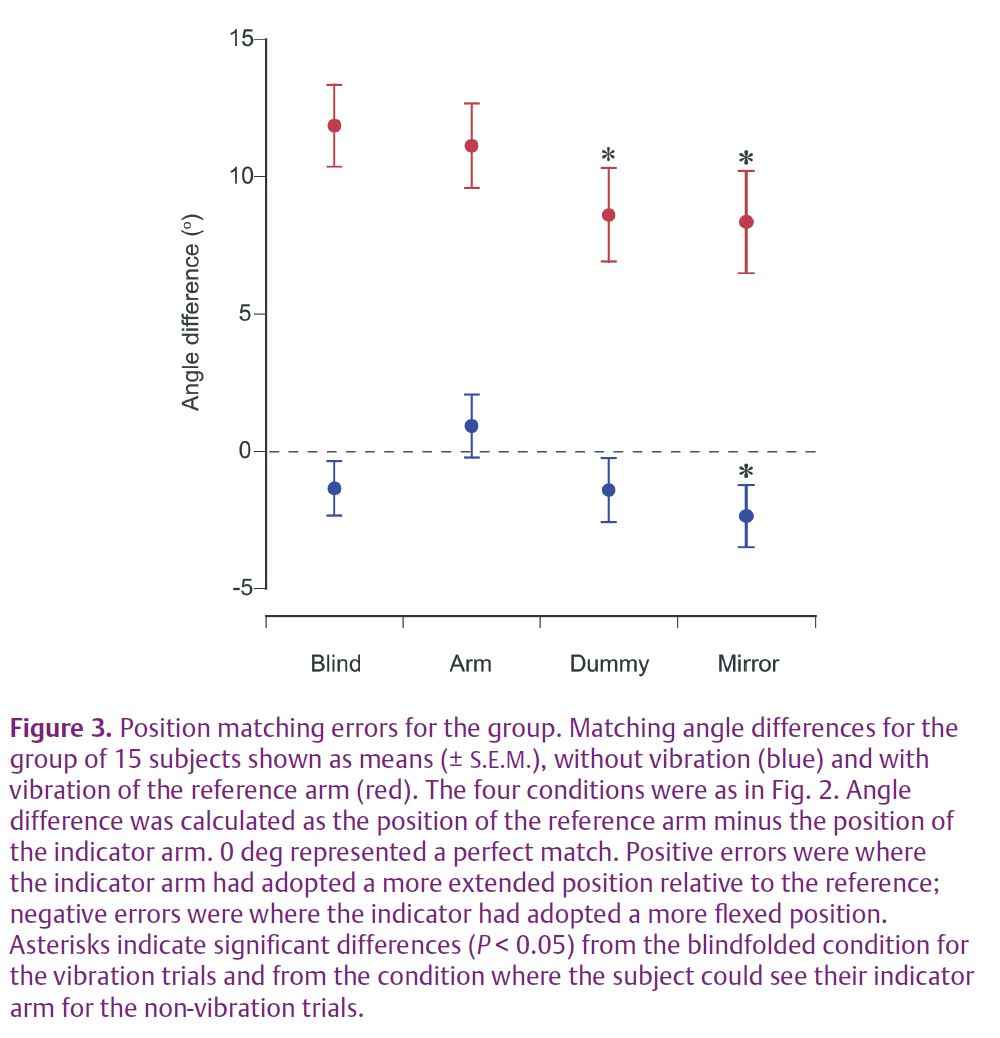
The first result was that for the matching series where the reference arm was not being vibrated, subjects managed to achieve comparable matching accuracy no matter what the condition of the indicator. Errors were similar (Fig. 3) when subjects were blindfolded and could not see either arm, or they could see the indicator or its mirror image, or they used the dummy to match positions. This last result is interesting in that it implies subjects were equally able to match position using proprioceptive information only (blindfolded) or visual information only (dummy). However, significant differences between conditions emerged when the reference arm was vibrated.
Vibration of the reference arm elicited a strong illusion that the arm was moving into extension and that its position was more extended than really was the case (Figs 2 and 3). The brain was falsely interpreting the muscle spindle signal from vibration as an elongating muscle, that is, an extending forearm. The size of the measured illusion was largest when the subject was blindfolded and they relied entirely on proprioceptive signals for matching. Here, presumably, the subject was trying to match the spindle signal coming from the non-vibrated arm with the much larger vibration-evoked signal coming from the reference arm. They did this by adopting an extended position with their indicator, in the process stretching its flexor muscles and raising the flexor spindle signal to a level somewhere near that coming from the reference arm. The observed error was 12 deg (Fig. 3).
Interestingly the error was only slightly smaller, 11 deg, when subjects matched while being able to see their indicator arm (Fig. 3). However, the errors were significantly smaller, 8.6 deg, when subjects used a dummy to match and 8.4 deg when they looked at a mirror image of their indicator arm during matching. With the dummy, subjects were using a visual cue to match the position of a proprioceptively perceived position.
In a simple interpretation, based on experience, vision of a particular arm posture is calibrated in terms of the normally expected proprioceptive signal. The discrepancy between the expected position indicated by sight of the dummy and that indicated by the proprioceptive signals coming from the reference arm led to a reduction in the position errors produced by the vibration. We think something similar was occurring when subjects looked at the mirror image of the indicator arm. Here an additional factor was likely to be that subjects were unsure of which arm they were looking at. In the trials where subjects simply looked at the indicator arm during matching, vibration errors were only slightly smaller than in the blindfolded condition as here both visual and proprioceptive signals coming from the indicator would be available and, very probably, the proprioceptive signal would be prioritised.
The main conclusion from this first set of experiments was that the condition of the indicator arm clearly played an important role in determining the accuracy of forearm position matching. Under normal circumstances, vision and proprioception can be used interchangeably during matching. However, if a false proprioceptive signal is introduced in one arm with vibration, this creates a conflict between the information provided by the proprioceptive and visual inputs from the two arms to reduce the effect of the vibration.
All of the observations described so far were concerned with the sense of forearm position. What about the sense of movement? The way we explored this was as follows: a vibration illusion was generated in one arm as before, but this time subjects were asked to track the perceived movement with the vibrated arm itself. So now during vibration the arm is slowly extending according to what the subject felt. We measured the speed of the movement on a computer. Throughout this time the other arm was held still at a particular angle. Then the experimenter began to move the non-vibrated arm in either the direction of extension or flexion. When the movement was in the direction of extension, subjects were seen to slow the speed of their tracking movement with the vibrated arm. When the movement was into flexion, movement of the vibrated arm speeded up (Fig. 4). Remarkably, when subjects moved their non-vibrated arm up and down in a continuous motion, they reported a ‘flutter’ sensation of slowing and speeding-up of their vibrated arm, even when this was not moving at all. There can be no more dramatic example of an interaction between proprioceptive signals coming from the two arms than this change in the quality of the sensation evoked by vibration in one arm that depended entirely on what the other arm was doing.
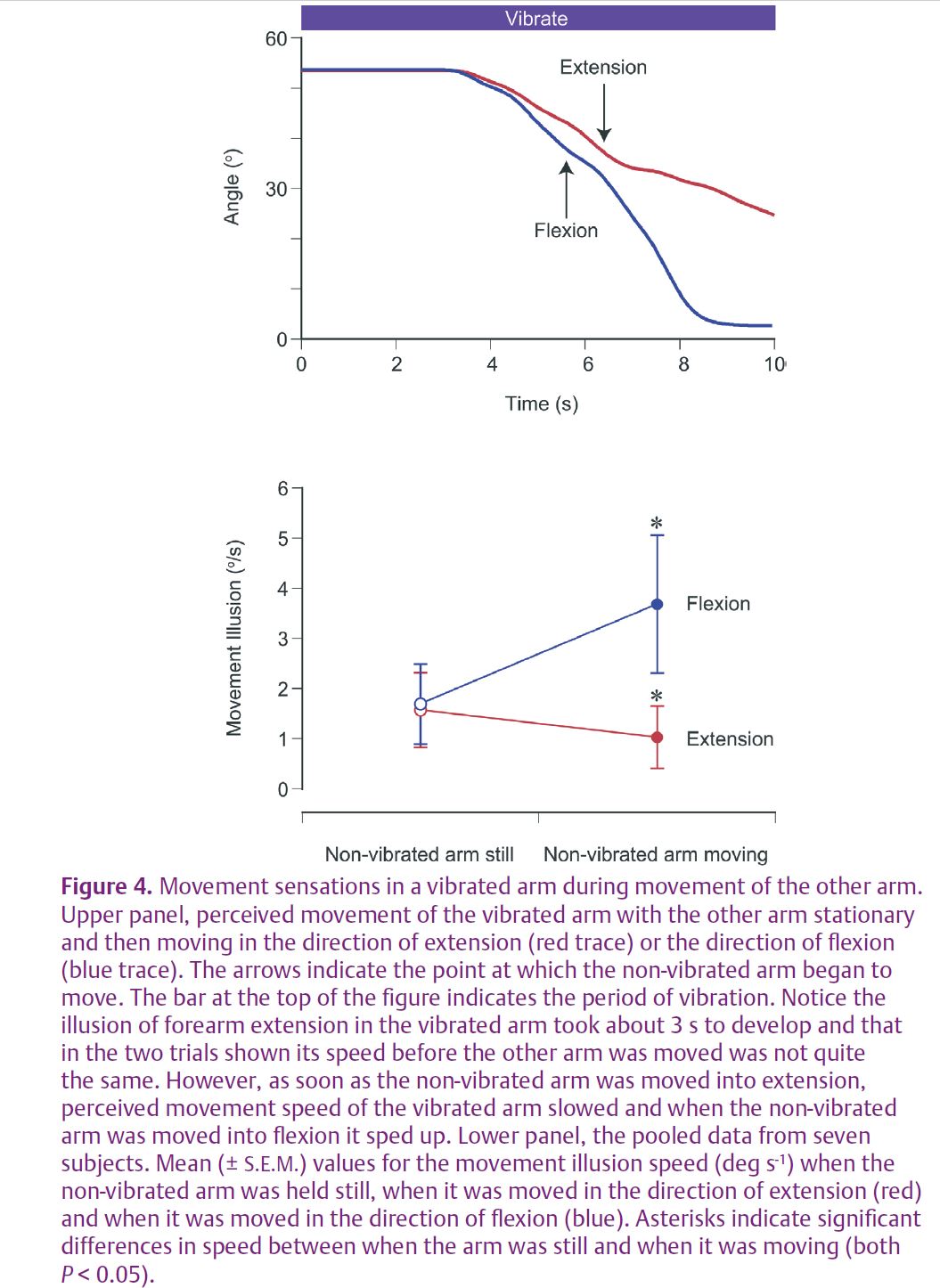
Our interpretation of this result is that what is felt during vibration is based, at least in part, on a difference signal derived from the inputs from the two arms. When the non-vibrated arm was moving into extension its elbow flexor muscles would be stretched and flexor spindles would therefore raise their level of discharge, thereby reducing the difference in proprioceptive signal between it and the vibrated arm. It led to slowing of the tracking movement by the vibrated arm. Conversely, if the non-vibrated arm was flexed, its spindles would be slackened by the muscle shortening and their discharge would be slowed, increasing the difference in signal between it and the vibrated arm. That led to an increase in speed of the tracking movement.
The present study has shown that both the sense of limb position and the sense of limb movement of one arm are dependent to some extent on the information coming from the other arm and that this information can be either visual or proprioceptive. Such a conclusion should be kept in mind in experiments on proprioception allegedly involving only one arm. Input from the other arm may not be entirely irrelevant. When the two arms are involved in a motor task where they are required to act cooperatively, for example, fine manipulations that require both hands close together, the central comparator is influenced by information coming from both arms, be it proprioceptive, visual or both. At the same time the identity of each arm is preserved. So in some circumstances the two arms can be considered to be acting as one, a single instrument, in the execution of certain skilled tasks.
References
Goodwin GM, McCloskey DI & Matthews PBC (1972). The contribution of muscle afferents to kinaesthesia shown by vibration induced illusions of movement and by the effects of paralysing joint afferents. Brain 95, 705–748.
Izumizaki M, Tsuge M, Akai L, Proske U & Homma I (2010). The illusion of changed position and movement from vibrating one arm is altered by vision or movement of the other arm. J Physiol 588, 2789–2800. http://jp.physoc.org/content/588/15/2789.long
McCloskey DI (1973). Differences between the senses of movement and position shown by the effects of loading and vibration of muscles in man. Brain Res 63, 119–131.
Proske U & Gandevia SC (2009). The kinaesthetic senses. J Physiol 587, 4139–4146.
Acknowledgement
We would like to thank Professor Ikuo Homma for his insight and encouragement during the project.