
Physiology News Magazine

Unravelling the processes that underlie the everyday activity of tracking moving objects
Features
Unravelling the processes that underlie the everyday activity of tracking moving objects
Features
Rochelle Ackerley (1,2) & Graham R. Barnes (2)
1: Faculty of Life Sciences, University of Manchester, Manchester, UK
2: Current address: Clinical Neuroscience & Department of Physiology, Sahlgrenska Academy, University of Gothenburg, Gothenburg, Sweden
https://doi.org/10.36866/pn.86.30
Specialised mechanisms have developed in the brain to allow us to follow moving objects. Our recent work has shown that exposure to only brief visual motion can be assimilated and used to guide eye and head tracking movements, even when an object disappears.
In our everyday lives, we follow moving objects automatically with our eyes. This is called smooth pursuit and it is difficult to generate smooth pursuit eye movements with no moving visual stimulus. When an object moves, we are quickly able to estimate its direction and speed to track it in space (Carl & Gellman, 1987). In addition, we generate an internal representation of the object’s velocity, from exposure to the motion combined with cognitive factors such as previous experience, prediction, expectation, learning and memory. This means we can continue tracking the object if it disappears from sight. In this situation, we are able to switch from using a combination of visual and internally driven feedback to produce the tracking response, to just using our internal representation. However, this tracking response can only be sustained if we can predict the movement trajectory of the object. If we expect the object to re-appear, smooth pursuit can continue for some time but if we believe it will not reappear, the tracking response decays (Becker & Fuchs, 1985).
For example, if we see a bird flying, it is easy to pursue when you have visual feedback. As well as the direct visual feedback, the pursuit response has an internally generated element so pursuit can become more automatic and you can do other things at the same time, like hold a conversation. This means we are using prediction to make assumptions about near-future events. If the bird flies behind a tree and disappears from sight, we can still continue to pursue its trajectory as we can match our eye velocities to the bird’s flying velocity by using only our internally generated response. This enables us to continue to move our eyes until the bird reappears from behind the obscuring object because there will be a reasonable expectation of when and where the bird will reappear. However, if the bird flies behind a large building, the expectation for future motion would be much lower and the tracking response would be more likely to decay. Recent work has shown that at least 100–150 ms of prior visual motion is required to drive pursuit when a moving target disappears (Barnes & Collins, 2008). In that paper, subjects either pursued a moving target with an expectation that it would disappear totally or reappear. In trials when subjects expected the target to reappear, smooth pursuit eye movements were continued at a velocity near to the velocity of the unseen target for up to 600 ms. These were guided solely by their internal velocity representation of the target. The internal velocity representation driving the eye pursuit was updated on a trial-by-trial basis, as eight randomised different velocities of target motion were used and the eye movements were scaled to the specific trial velocity.
The situation is more complicated when we track a moving object with our eyes and head together. When we rotate our heads, the vestibulo-ocular reflex (VOR) is invoked, which drives the eyes in the opposite direction at a near-equal rate to the head rotation (for a review of vestibular mechanisms, see Angelaki & Cullen, 2008). This fast reflex allows us to maintain fixation on an object when our head moves. However, if we want to follow a moving object with our eyes and head together, we need to countermand this reflex to move our eyes and heads in the same direction. The mechanisms that allow this countermanding to take place are principally the same as those that control pursuit eye movements, although other non-pursuit mechanisms are thought to operate in some circumstances. But previous attempts to examine the role of pursuit in this context have not been able to segregate the visually driven and internally driven components and it was with this in mind that we (Ackerley & Barnes, 2011) investigated the mechanisms involved in the pursuit of a moving target that disappeared from view at specific times, using the eyes and head together.
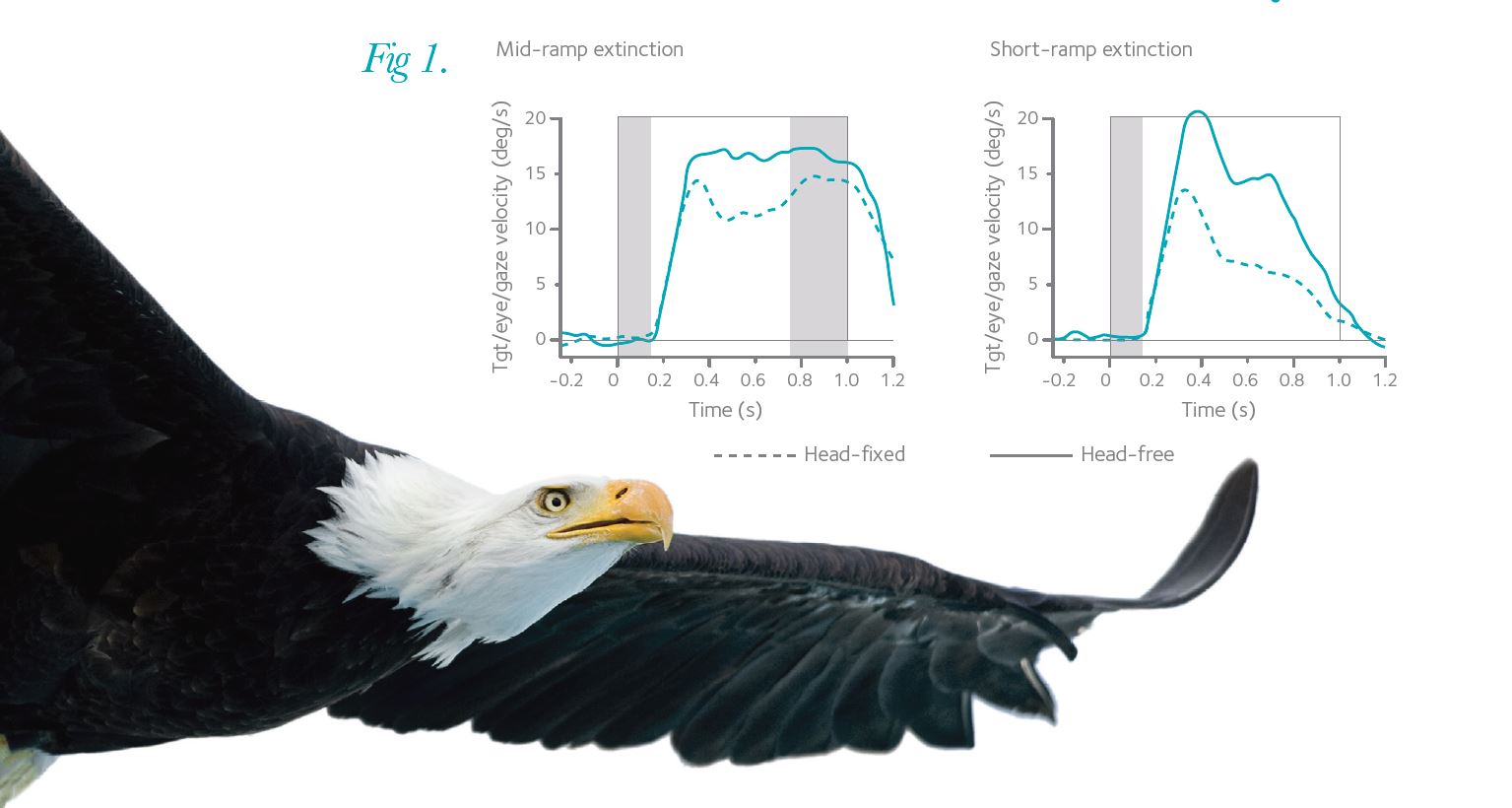
This series of experiments was designed to uncover features of the internal pursuit response under more natural circumstances. The experiments used a novel protocol that allowed the visual and internal components of pursuit to be segregated, during head-fixed (with eyes only) and head-free (eyes and head) pursuit. In the experiments, the subjects viewed the beginning and/or the end of the visual motion trajectory, using different, randomised velocities, and were instructed to try and pursue the moving target when it disappeared. In experiments where the subjects viewed the first 150 ms and the final 200 ms of visual motion, with an intervening 600 ms period of target extinction, they were able to continue eye or gaze (eye + head) movements during the period of unseen, but predictable motion (Fig. 1, mid-ramp extinction). In contrast, when the subject viewed only the first 150 ms of visual motion and had no expectation that the target would reappear at a later point, eye and gaze velocity decayed after an initial tracking response (Fig. 1, short-ramp extinction). During pursuit using the head, the subjects were able to obtain a higher pursuit velocity throughout periods where the target was unseen than in trials when they only moved their eyes.
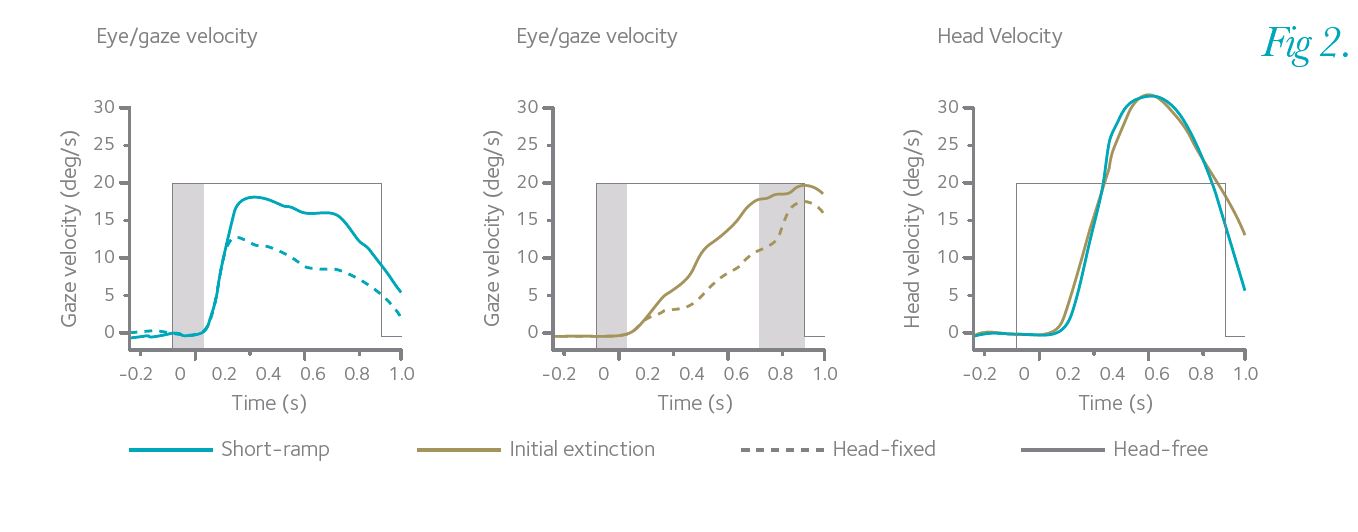
In further experiments, the mid-ramp extinction condition was split into pairs of identical randomized velocity stimuli, where the first presentation was 150 ms motion (equivalent to the short-ramp extinction); it was followed by a short break where the target was stationary in the centre of the screen. In the second of the pair, the target disappeared from the centre, which signalled the onset of (unseen) target motion and only the last 200 ms motion was seen (initial extinction). Subjects had to use the motion information in the first of the pair to make an attempt to pursue the second of the pair.
Figure 2 shows that subjects were, indeed, able to successfully make smooth pursuit movements using only the previously remembered motion. This pursuit to the second stimulus was solely driven by internal mechanisms and showed that the brain was using expectations about the future target movement to predict the trajectory, thus doing so with no direct visual motion. Again, the gaze response velocity was higher in the head-free condition than when the head was fixed.
The results show the internal mechanisms for pursuit are similar in head-fixed and head-free conditions. This internal drive is also able to countermand the effects of the VOR, even under conditions when there is no visual stimulus. Detailed analysis revealed that the VOR itself remained active throughout the period of target disappearance, but that, because its gain was slightly less than unity, it resulted in the observed increase in gaze velocity in the head-free condition. In summary, the findings support the view that cognitive mechanisms, such as expectation, can modify smooth pursuit when objects disappear from sight. Furthermore, if we use our heads to attempt to track an unseen object, we can achieve better pursuit, which is of relevance to wider areas, for instance clinical rehabilitation strategies. All of this reveals that the brain is constantly using internal predictions to aid us in our everyday lives, rather than having to rely on sensory feedback, which has an innate delay.
Acknowledgments
The work from the authors was supported by a grant from the Medical Research Council, UK.
