
We have demonstrated that the temporal discharge patterns of neurons in ferret primary auditory cortex (A1) transmit significant amounts of information about stimulus identity when tested with natural and time-reversed marmoset ‘twitter’ vocalisations (Schnupp et al. 2006). We have compared the discrimination performance afforded by these spike pattern codes (a ‘neurometric’ function) to behavioural performance (a psychometric function). Psychophysical data were collected for a two-alternative forced-choice oddity task in which 6 human observers had to distinguish natural twitter recordings from those with local time reversals of 10, 20, 40 or 80 ms width (‘flipped twitters’). Listeners’ discrimination performance was near perfect when the reversed time windows were 80 ms wide, but declined dramatically for time windows of 20 ms or less. We also recorded responses of 142 A1 neurons to these stimuli in 3 adult ferrets. Anaesthesia was induced by 2 ml/kg intramuscular injection of alphaxalone/alphadolone acetate, and was maintained with intravenous infusions of medetomidine/ketamine at a typical rate of 0.022 and 5.0 mg/kg/h, respectively (as described in Garcia-Lazaro et al. 2006). We used methods derived from signal detection theory (Green & Swets, 1974) to determine if the temporal discharge patterns of A1 responses could discriminate between the natural and flipped twitters. While no individual unit’s neurometric matched the psychometric performance curve perfectly (Fig. 1), the neurometric of enveloped or pooled responses of the population of units in our sample closely resembled the psychometric curve (Fig. 2). Therefore, the statistical properties of temporal discharge patterns of distributed populations of A1 neurons show similar stimulus-related effects to the behavioural discrimination of complex stimuli, suggesting that this neural code may underlie the percept of the stimulus. However, neurometrics based on overall spike counts could not account for behavioural performance. Neurometrics performed best when A1 temporal discharge patterns were analysed at a resolution of less than 10-20 ms.
University College London 2006 (2006) Proc Physiol Soc 3, C96
Research Symposium: A neurometric analysis of spike pattern codes for natural and manipulated vocalization stimuli in primary auditory cortex
Kerry Marie May Walker1, Andrew J King1, Bashir Ahmed1, Jan W H Schnupp1
1. Physiology, Anatomy & Genetics, University of Oxford, Oxford, United Kingdom.
View other abstracts by:
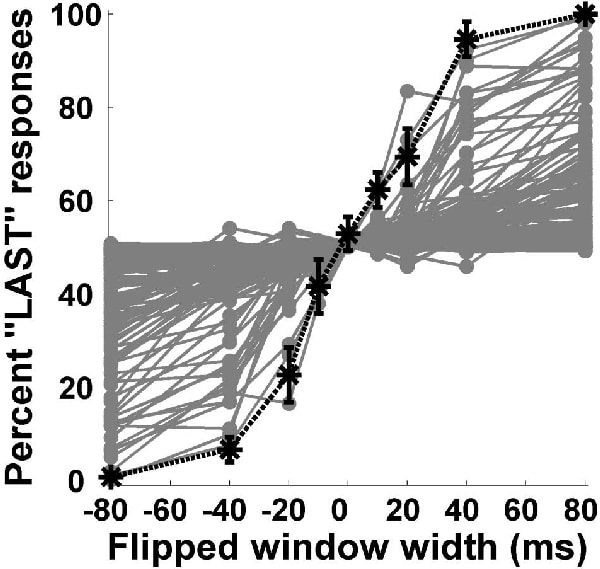
Figure 1. Neurometric functions for all 142 units in our sample are plotted superimposed (grey circles) along with the average psychometric performance (mean ± SEM) of the human listeners on the 2-alternative forced choice (2AFC) task (black asterisks). Negative 'Flipped window widths' indicate that the oddball stimulus was the first of the 3 stimuli presented in the 2AFC trial while positive values indicate that the oddball stimulus was last in the sequence.
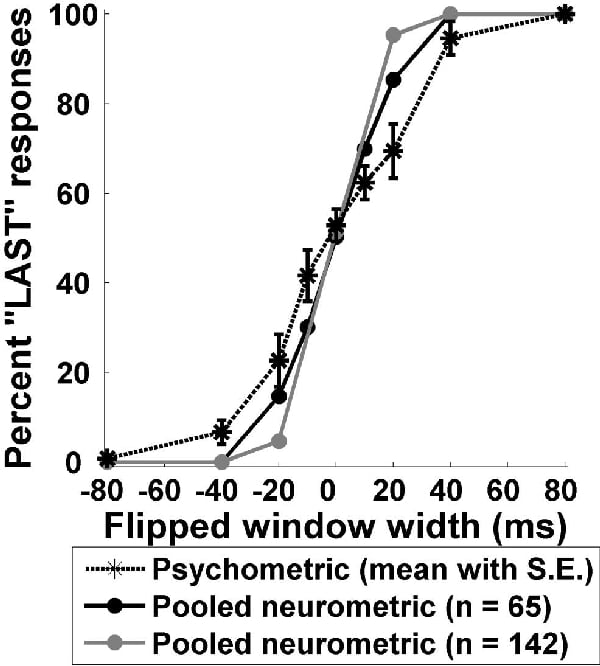
Figure 2. Population neurometrics obtained from the pooled responses of 65 (black circles) and 142 (grey circles) units are shown along with the human psychometric (mean ± SEM) curve (black asterisks).
Where applicable, experiments conform with Society ethical requirements.