
Physiology News Magazine

The sodium hypothesis of action potential generation
A classic updated or an act of electrophysiological sacrilege?
Features
The sodium hypothesis of action potential generation
A classic updated or an act of electrophysiological sacrilege?
Features
Angus Brown
University of Nottingham, UK
https://doi.org/10.36866/pn.114.32
Read the Journal of Physiology virtual issue here, or updated Hodgkin and Katz paper as a PDF here.
In 1972, Stephen Kuffler asserted students should consult original sources (Kuffler, 1972), a viewpoint with which I concur. By the time experimental information has reached textbooks, the inevitable dilution, interpretation and editing may obscure the researchers’ original intentions. For neuroscience students interested in membrane excitability there is a rich history of such information, a considerable amount of which is freely available via The Journal of Physiology. However, there is one major impediment that stands in the way of students’ appreciation of these papers: the conventions used to present and describe data differ radically from those used today, which can be confusing even for the experienced researcher.
In an effort to surmount this I have dared to update one of the founding classics of electrophysiology, namely “The effect of sodium ions on the electrical activity of the giant axon of the squid” (Hodgkin and Katz, 1949). The difficulty in reading this paper stems from Hodgkin and Katz using the accepted norm in physics that an inward movement of positive charge is depicted as an upward current deflection, with the resulting change in voltage depicted as a downward deflection (Patton, 1982). Hodgkin and Katz treated the resting membrane potential as a positive quantity and the action potential (AP) as a negative deflection; thus, the action potential was displayed as an upward deflection with the y-axis defined as negative volts (–V).
Modern practice deviates from this convention, showing inward current as a downward deflection and membrane potential as “positive-up, negative-down” (Patton, 1982). The modern convention is illustrated in Fig. 1, where an electrode is shown inserted into a cell submerged in a bath, the amplifier is grounded (0 mV), and a resting membrane potential of about -70 mV is recorded, leading to the classic physiology teacher’s formulation where, if the membrane potential (Em) moves towards 0 mV this is considered a depolarisation (a decrease in polarisation), a move towards the resting potential is considered a repolarisation, and a move to a potential more negative than the resting potential is considered a hyperpolarisation (an increase in polarisation; Hille, 2001).
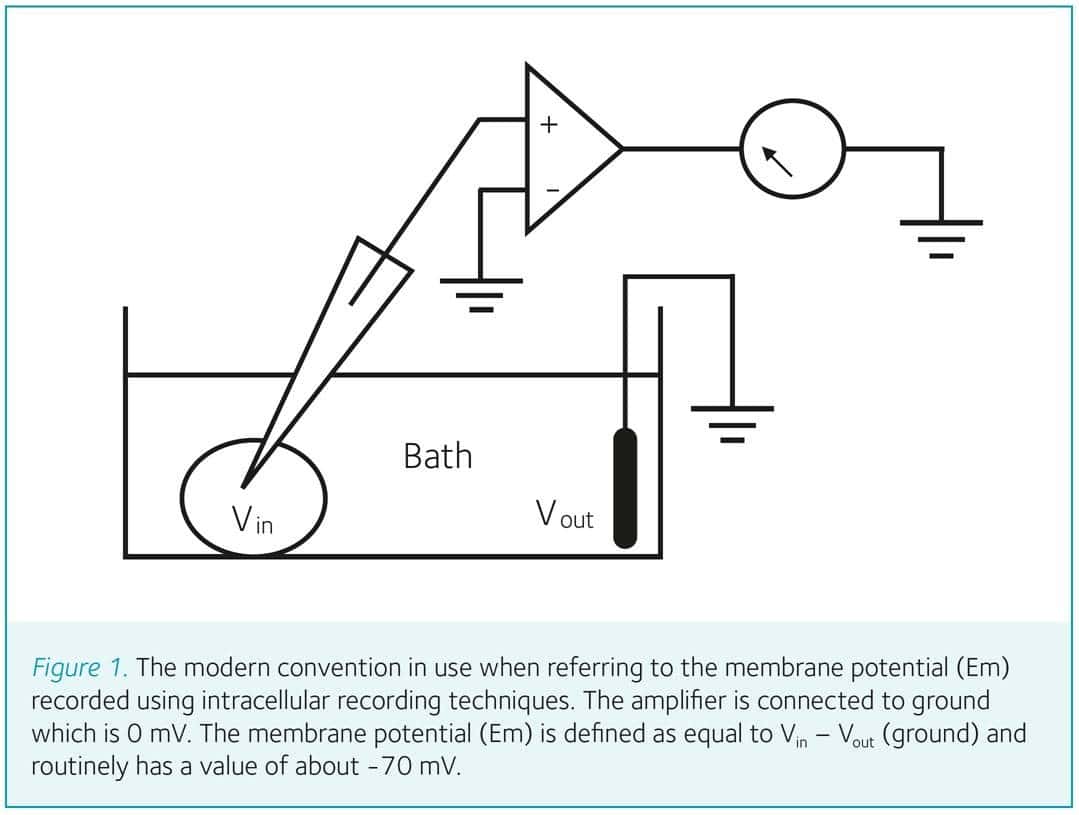
The convention used by Hodgkin and Katz is based upon a reference system where changes in Em were considered relative to rest, with increases or decreases, respectively, being equivalent to hyperpolarisation or depolarisation, respectively. This was due to technical considerations, since the junction potentials created by switching from seawater to the test solutions made accurate measurements of the potential difference across the axon membrane unreliable. Indeed, Hodgkin and Katz spent a considerable amount of the paper discussing junction potentials and their effects on Em. A source of confusion is that the manner in which Hodgkin and Katz displayed the Em was inconsistent. Although resting potential is shown as +45 mV in Fig. 4 of the original paper, in Fig. 3 it is shown as 0 mV with the action potential shown as a positive voltage deflection, and in Fig. 5 both the resting potential and the action potential are shown as positive values. This confusion is compounded when deviations of Em from rest are variously described as “depression, decrease, diminution, reduce, lowered, raising, larger”, each description being context-dependent. In addition, some of the terminology in the paper is antiquated e.g. the “positive phase” refers to the after hyperpolarisation (AHP), and “membrane reversal” refers to the degree by which the action potential exceeds 0 mV, i.e. overshoot, terms that are now redundant. The differences between the historic and modern convention are illustrated in Fig. 2.
The history of the paper’s origin is captivatingly described in detail elsewhere (McComas, 2011), but may be briefly summarised as follows. The dogmatic view, proposed by Bernstein at the beginning of the 20th century (Hille, 2001), was that the resting neuronal membrane is predominantly permeable to K+, but this selective permeability collapses during an action potential. This theory predicted that Em should approach 0 mV at the peak of an action potential. However, the first intracellularly recorded action potential, obtained from squid giant axon (Hodgkin and Huxley, 1939), famously showed a peak that exceeded 0 mV and approached ENa. It is often forgotten that Hodgkin and Huxley made no reference to ENa in the original paper, or indeed in a later more extended account (Hodgkin and Huxley, 1945), but instead provided four possible explanations for the membrane reversal, none of which mentioned Na+ permeability.

Their reluctance to make the obvious connection between the action potential and ENa lay in a paper published by Curtis and Cole in 1942, in which the authors described action potentials whose peaks occurred at potentials more positive than ENa and, more worryingly, showed that removal of Na+ from the bathing solution did not significantly affect the AP amplitude (Curtis and Cole, 1942). Despite this, Bernard Katz suspected an important role for Na+ in action potential generation, and showed in 1947 that decreasing bathing [Na+] decreased the conduction velocity in extracellularly recorded action potentials in single fibres of the crab Carcinus (Katz, 1947). Hodgkin was subsequently joined by Katz at Plymouth in the summer of 1947 (Huxley was busy preparing for his wedding), where they quickly and satisfactorily resolved the issue by recording the Em with intracellular microelectrodes in squid giant axon exposed to various dilutions of seawater, such that the data was acquired by September 1947 and submitted for publication in January 1948.
Although the main interest in the paper was the effect of [Na+] on the action potential, the effect of [K+] on the resting potential was of equivalent importance although of a less dramatic nature. The main findings of the paper were:
- AP amplitude was dependent upon [Na+]
- Rate of AP rise was dependent upon [Na+]
- Conduction velocity decreased in low [Na+]
- Rate of AP fall was independent of [Na+]
- Resting Em was dependent upon [K+]
- After hyperpolarisation amplitude was dependent upon [K+]
- The permeability changes underlying the AP could be quantified using the Goldman–Hodgkin–Katz equation, an extension of the Nernst equation, and supported a transient increase in Na+ permeability underlying the AP
From an historical viewpoint, the paper is the foundation from which flowed the subsequent papers that gained Hodgkin and Huxley the Nobel Prize in 1963. In hindsight, the paper is not above criticism. It is rather verbose and repetitive by modern standards, and could easily be condensed to one third of its original length without losing any impact. The authors are circumspect in describing the now obvious relationship between Em and reversal potentials for Na+ and K+, and only in the discussion do they tentatively speculate on this subject, likely due to uncertainties of the values of the internal ion concentrations. For this reason, in appropriate figures, I have judiciously added reversal potentials for K+ and Na+ for illumination. It is notable that the resting membrane potential recorded by Hodgkin and Katz was depolarised compared to values recorded later with less intrusive methods (-45 mV compared to -70 mV; in Chang, 1986). This resulted in the resting potential being between 40 and 50 mV more depolarised than EK, at odds with the central tenet proposed by Bernstein, and expounded by Hodgkin and Katz, of the membrane being selectively permeable to K+ at rest.
Reading the complete paper evokes an unmistakable sense of place and time – a time when the unfolding of a scientific story included leisurely discourses. The paper is without doubt a classic, my personal favourite in the entire electrophysiology canon. Updating it has been a labour of love, not a chore (though some might consider it more of a sacrilege!). The logical sequence of Hodgkin and Katz’s figures is masterly at revealing critical data that is described later. For example, a glance at Fig. 3 reveals that the AP falls on exposure to Na+-free seawater. However, hiding in plain sight is the fact that the falling phase of the AP, the AHP and the resting potential are all relatively unaffected by changes in external Na+. In Fig. 8, one of the most reproduced images in all of neuroscience, the AP amplitude is shown to increase in axons exposed to supra-physiological concentrations of Na+ – simple, brilliant and entirely convincing evidence that the peak amplitude of the AP follows ENa.
I will not list all the changes I have made to the paper, but will only state that they comply with modern convention and should be readily understood by all students raised on current textbooks. I have inserted equations that illuminate calculations in the text, included only averaged data in the Tables, added explanatory columns in Table 4 relating to ratios of [Na+] relative to seawater, and redrawn graphs where appropriate. A thorough understanding of the practical implementation of the Nernst equation is recommended for optimal appreciation of the paper (Sawyer et al., 2017). I have omitted the Appendix on the derivation of the equations, as this does not require extensive conversion and is of limited interest to the majority of undergraduate students.
In conclusion, the classic electrophysiology literature of the 1940s and 1950s is at risk of becoming a museum piece; admired and respected, but ultimately inaccessible, as the contemporary scientists age and no longer pass their knowledge on to succeeding generations of students. It would be a terrible shame if this were to occur, but hopefully this article may tempt qualified and dedicated individuals, with sufficient spare time, to proceed with equivalent updates of their favourite classic papers.
References
Chang DC (1986). Is the K permeability of the resting membrane controlled by the excitable K channel? Biophysical Journal 50(6), p.1095–1100.
Curtis HJ, Cole KS (1942). Membrane resting and action potentials from the squid giant axon. Journal of Cellular and Comparative Physiology 19, 135–144.
Hille B (2001). Ion Channels of Excitable Membranes. Sunderland: Sinauer Associates Inc.
Hodgkin AL, Huxley A (1939). Action potentials recorded from inside a nerve fibre. Nature 144, 710–711.
Hodgkin AL, Huxley A (1945). Resting and action potentials in single nerve fibres. J Physiol 104, 176–195.
Hodgkin AL, Katz B (1949). The effect of sodium ions on the electrical activity of the giant axon of the squid. J Physiol 108, 37–77.
Katz B (1947). The effect of electrolyte deficiency on the rate of conduction in a single nerve fibre. J Physiol 106, 411–417.
Kuffler SW (1972). Introduction. In: Cellular Neurophysiology, ed. Cooke I & Likin M, pp. 1–3. Holt, Reinhart & Winston, Inc., New York.
McComas AJ (2011). Galvani’s Spark. Oxford: Oxford University Press.
Patton HD (1982). Special properties of nerve trunks and tracts. In Physiology and Biophysics: IV Excitable Tissues and Reflex Control of Muscle, ed. Ruch T & Patton H, pp. 101–127. W.B. Saunders Company, Philadelphia.
Sawyer JER et al. (2017). The critical role of logarithmic transformation in Nernstian equilibrium potential calculations. Adv Physiol Edu 41, 231–238.